Dit blog is een onderdeel van EVODISKU WAT IS DE BEDOELING EN WAAR STAAT DIT BLOG VOOR
****Wie meent dat alles inmiddels wel over de evolutietheorie gezegd is en dat de discussie gesloten kan worden, ziet over het hoofd dat de wetenschap niet stil staat.
***Wie meent dat inmiddels het creationisme definitief het pleit heeft verloren en dat de discussie gesloten kan worden , ziet over het hoofd dat het "creationisme" is geevolueerd ( en zal evolueren ) in nieuwere mimicrytische vormen( meme-complexen ) zoals bijvoorbeeld het ID(C)
***Dit blog is speciaal opgezet om de aktualiteit binnen het evolutie-creationisme debat te volgen en van kommentaren te voorzien ... waartoe de lezers zijn uitgenodigd bij te dragen ...
Let echter wél op het volgende :
"Je bent een rund als je hier met religie stunt " ....
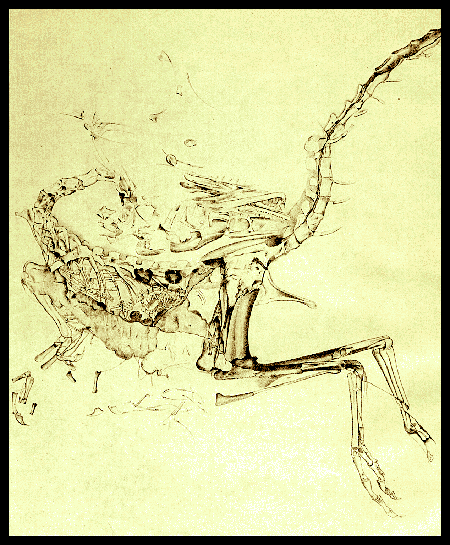
Chinese onderzoekers( = paleontologen van de Chinese Academie van Wetenschappen in Bejing met o.a.Fucheng Zhang & all ) hebben een vreemdsoortige theropodedinosauriër, van de Maniraptora, groep beschreven ; Het holotype IVPP V15471, van de Epidexipteryxhui (1) is in het binnenland van Mongolië gevonden eind 2007 (Daohugou Beds ) Het lichaam van het beest was wel bedekt met veerachtige structuren , maar heeft nooit kunnen vliegen. Dat staat in een artikel dat in het Britse wetenschapstijdschrift Nature gepubliceerd op 23 oktober 2008
-Het Epidexipteryx hui fossiel gevonden in Mongolia bezit lange stijve staartveren terwijl het lichaamis bedekt met kort "dinodons " Deze vondst vertegenwqoordigd ;de oudst bekende verwant van de vogels en de maniraptora Het gebruikte de veren misschien meer voor de balts en het pronken dan om te vliegen ...althans dat werd in een September 2008 paper, al geopperd foto: Fucheng Zhang et al.
Als de datering van de vogel klopt, is hij ouder dan het beroemde vogelfossiel Archaeopteryx en ouder dan de vele fossielen van vogelachtige dinosauriërs die ook in China zijn gevonden.
Pronken
Epidexipteryx is Grieks voor 'baltsveren'. Het beest bezat een korte staart maar was getooid met vier opvallend lange pauwenveren, vermoedelijk om mee te pronken tijdens de balts De baarden van die veren ontbreken in het fossiel
Verder was zijn huid bedekt met een laag veren: deze lichaams-bedekking , het bij maniraptora gekende "dinodons" (2) , heeft geen bewaarde pennenschachten ,slechts paralelle baardjes .... Bij het Epidexipteryx fossiel echter lijken de "veren" aan de basis een continu membraan te vormen ; pas aan de uiteinden eindigen ze in parallelle "draadachtige " stuctuurtjes .Deze eigenschap is hoogstwaarschijnlijk uniek Er zijn bovendien ook geen slagpennen op de armen gevonden Anders dan andere Chinese vogeldinos als Microraptor heeft Epidexipteryx geen slagpennen op de armen . Dit zijn stevige veren die een vogel nodig heeft om te vliegen. Het is voornamelijk om die reden en het feit dat het weliswaar relatief lange armen bezat maar wel ledematen die te kort zijn om een voldoende spanwijdte te kunnen verwezenlijken , niet waarschijnlijk dat de Epidexipteryx het luchtruim kon kiezen .....
Het is ook mogelijk dat het een lopend dier betreft dat afstamt van een vroegere vlieger ; vooral de klauw aan de voorarm zou dat suggereren (2) Maar het dier zou wel een dicht verwante voorloper van de Archaeopteryxkunnen zijn
Wetenschappers denken dat de Epidexipteryxin elk geval een representatief puzzelstukje is uit de overgang. naar de Avialae, en het is daarbij (morfologisch) nauw verwant aan de in dezelfde formatie gevonden Epidendrosaurus, binnen de Scansoriopterygidae. : Omdat hij geologisch ouder is dan de Archaeopteryx staat hij waarschijnlijk ook dichter bij de maniraptora dan de vogels
De Epidexipteryx leefde ergens +160/150 miljoen jaar geleden( er bestaat nog geen consensus over de datering ) , in het midden- tot laat Jura-tijdperk. Als de datering van de vogel klopt, is hij ouder dan het beroemde vogelfossiel Archaeopteryx en ouder dan de vele fossielen van vogelachtige dinosauriërs die ook in China zijn gevonden. Het was maar een klein schepsel; zo groot als een duif en woog slechts 160 gram. Het dier had een langwerpig hoofd en opvallend lange voortanden.
FIG 1= Epidexipteryx hui gen. et sp. nov., IVPP V15471, main slab and close-up photos
a, Main slab; b, c, skull in main slab (b) and counterslab (c); d, four elongate ribbon-like tail feathers; b', c', line drawings of b and c, respectively. Abbreviations: l1, l2 and l7, 1st, 2nd and 7th left teeth of upper jaw; l1', r1' and r5', 1st left, 1st right and 5th right teeth of lower jaw; l2 and r2, 2nd left and right teeth of upper jaw.
FIGURE 2=Line drawings and close-up photographs of Epidexipteryx hui gen. et sp. nov
a, Skeleton and feather outline, based on both main slab and counterslab, showing that each shafted feather is formed by central rachis and two unbranched vanes (a'). b, d, d', Proximal regions of non-shafted feathers; barbs are parallel and closely united as an unbranched membranous structure (d, d'), vanes are either layered, indicated by white arrows (b), or arranged irregularly (c). The red arrow indicates the proximal end of the feather (d). e, e', f, Distal regions of non-shafted feathers, in which barbs appear loosely parallel. Abbreviations: as+ca, astragalus and calcaneum; ce, cervical vertebrae; co, coracoid; cv, caudal vertebrae; dt3+4, distal tarsals 3 and 4; f, frontal; fe, femur; fi, fibula; ga, gastralia; hu, humerus; il, ilium; is, ischium; ma, mandible; mtIIV, metatarsals IIV; p, phalanges or metacarpals; pa, parietal; pm, premaxilla; pu, pubis; q, quadrate; r, ribs; ra, radius; rc, rachis; sc, scapula; st, sternum; ti, tibia; tv, thoracic vertebrae; u, ungual phalanges; ul, ulna; uv, unbranched vane.
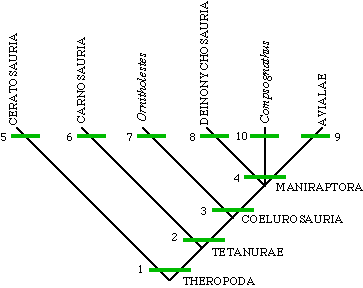
FIGURE 3 =Phylogenetic relationships of Epidexipteryx hui gen. et sp. nov.
The cladogram is simplified from the strict consensus of nine most parsimonious trees (tree length 1,255; consistency index 0.35; retention index 0.75; see Supplementary Information). Scansoriopterygidae is defined as the least inclusive clade including Epidendrosaurus and Epidexipteryx, Avialae as the most inclusive clade including Vultur gryphus but not Deinonychus antirrhopus, and Aves as the least inclusive clade including Archaeopteryx and Vultur gryphus.
Het dier vertoont een hoogst merkwaardige mix ( mozaiek? ) van kenmerken te vinden bij verschillende theropodegroepen ( vooral bij de Oviraptorosauria ) (3)....zoals in het huidige 23 oktober artikel onder de aandacht wordt gebracht
"This new species is characterized by an unexpected combination of characters seen in several different theropod groups, particularly the Oviraptorosauria. Phylogenetic analysis shows it to be the sister taxon to Epidendrosaurus4, 5, forming a new clade at the base of Avialae6. "
(2) Omdat het fossiel zeer goed bewaard is gebleven, is Zhang er vrij zeker van dat Epidexipteryx nooit slagpennen heeft gehad. Wel is het mogelijk, schrijft hij, dat de voorouders van Epidexipteryx konden vliegen en dat de dinovogel dit vermogen verloren had, net als de moderne struisvogel.
De andere verklaring voor de afwezigheid van de slagpennen is veel interessanter: Epidexipteryx en zijn voorouders konden niet vliegen. In de loop van de evolutie ontwikkelden zij veren om indruk te maken op soortgenoten zoals potentiële partners of rivalen. Later kwamen die veren ook van pas om te vliegen.
Of vogels hun eerste veren gebruikten om te vliegen, is een vraag die evolutionair biologen al lang bezighoudt. Een probleem is dat evolutionaire veranderingen stap voor stap verlopen, terwijl een functionele vleugel met veren een complex geheel is. Sommige biologen gaan ervan uit dat een paar eenvoudige veren dinosaurussen hielpen om een glijvlucht te maken vanuit een boom of om even van de grond te wippen. Een alternatieve verklaring is dat (dons)veren de dieren beter warm hielden dan haren. (3) Ik voorspel dat sommige Creationisten allicht zullen proberen deze Epidexipteryxhui ( vooral vanwege het mozaik- karakter van de anatomie ) in een kwaad daglicht te stellen.... en dat door vooral heel tendentieus en met halve waarheden te herinneren aan Archaeoraptor (de "piltdown kip" in het creationisten jargon ) , indien het al niet af te doen als een nieuwe fraude ?
Wat overigens niet wil zeggen dat de autenthiciteit van het fossiel NIET in vraag zou mogen worden gesteld...integendeel ; Dergelijke autentieke fossielen zijn van grote wetenschappelijke waarde ...maar zijn dat ook vanwege de hoge prijzen die daarvoor op de officiele verzamelaars - en ( vooral ) zwarte markt betaald worden ......
Archearaptor is wel degelijk een onecht fossiel in die zin dat het bestaat uit minstens ( zo werd in het begin geopperd toen de "vervalsing " werd ontdekt )drie aan elkaar geplakte fossielen van afzonderlijke soorten ( een chimaera, dus )
nml : De staart kwam van een gespleten fossiel van een kleine dromaeosauriër; de (gespleten stukken van de) voet van een nog onbekend dier en voor het linkerdijbeen en de onderbenen waren ook weer andere fossielen gebruikt....
Het staartfossiel werd uiteindelijk beschreven als Microraptor, de bovenkant (IVPP V12444) werd eerst Archaeovolans repatriatus genoemd ("oude vlieger die naar zijn land is teruggekeerd"), maar bleek in 2002 identiek te zijn aan Yanornis martini.
Het zijn bovendien soorten die behoren tot de verwante afstammelingen van de voorouders of vroege leden van de vogels uit dezelfde periode en afkomstig van dezelfde vindplaats .....
Microraptor is misschien in technische zin geen vogel, maar was desalniettemin een gevleugelde vorm die door iedere leek voor een "vogel" aangezien zou worden. Was Archaeopteryx niet bekend geweest dan zou Microraptor perfect de rol van dé oervogel hebben kunnen vervullen. Net zoals sommige echte vogels wel degelijk een lange staart hadden, zo hadden iets primitievere andere dinosauriërs wel degelijk vleugels.
Yanornis martini,[1] , een vroege vogel uit het krijt is beschreven aan de hand van een 5-tal fossielen gevonden in de Jiufotang Formation Chaoyang, /Westen van de Liaoning province, Het gedeelte dat is gebruikt in " archeoraptor" is dus ____net als het microraptor deel _____ een welkom zeldzaam fossiel
Compsognathus longipes is a small theropod, coelurosaurian dinosaur from Solnhofen. The type specimen (below) has in its gut region the remains of a small long-tailed lizard. There are no signs of scale or feather impressions. If a specimen such as this were to have impressions of its integument, there would be some basis for knowing if small dinosaurs has an insulatory coating of feathers.
On the right is a lithograph of Compsognathus longipes from Solnhofen. This lithograph was made on the lithographic limestone from Solnhofen itself. Such lithographic plates are the reason the quarries were opened in the first place. If you click on the image, you will go to a web site in which Compsognathus is described in more detail.
Maniraptora
1. three-toed foot 2. digits IV and V lost on hand 3. long arms 4. semilunate carpal 5. fused pelvis 6. large hole in lacrimal bone in skull 7. ?no unique derived characters? 8. giant, hook-like claw on digit II of pes 9. flight feathers 10. ?no unique derived characters?
Cladogram of the Theropoda
Aves
Archaeopteryx lithographica Below is the Berlin specimen of Archaeopteryx from the Solnhofen Limestone. It was the third specimen found, and unquestionably the most spectacular specimen found to date. Particularly noteworthy are the very fine feather impressions, including the flight feathers of the wings. Despite its' obvious avian nature, Archaeopteryx has a hand virtually identical to other maniraptoran dinosaurs such as Velociraptor.
Bird and Dinosaur Features
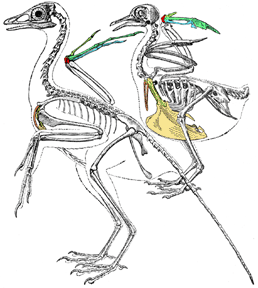
Comparison of reconstructed skeleton of Archaeopteryx (left) and a pigeon (right). Key to homologous bones follows: yellow, sternum (breastbone); orange, furcula = fused clavicles (wishbone); yellow, coracoids; red, semilunate carpal; yellow-green, digit I of manus; green, digit II of manus; blue, digit III of manus. This drawing is modified from Gerhard Heilmanns classic monograph "The Origin of Birds".
Note the long tail, separate fingures of the manus, and small sternum of Archaeopteryx, all primitive features inhereted virtually without modification of non-bird maniraptorans.
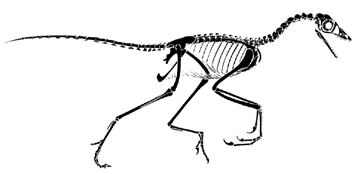
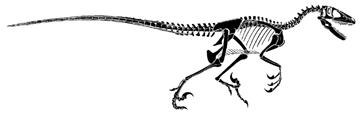
Reconstruction of Archaeopteryx (based on Paul, 1988). Note the amazing similarity to the sketon of another. larger maniraptoran, Deinonychus from the Early Cretaceous Cloverly Formation of Montana.
Reconstruction of the skeleton of Deinonychus (modified from Paul, 1987)
Some of the strongest and most convincing evidence of the maniraptoran origin of birds is in the comparision between the manus of various non-bird dinosaurs and a modern bird, the Hoatzin, from South America.
It is unique in a number of ways among birds. First, the young (see below) have claws of well developed fingers. They jump from their nests to escap predators (mostly snakes) and then return to the nest by clibing back up using their manus. In addition the Hoatzin is the only bird that has developed a foregut in which cellulose digestion takes place, aided by bacterial fermentation. It is a true folivore - it eats leaves (see www.knowledge.co.uk/frontiers/sf066/sf066b06.htm).
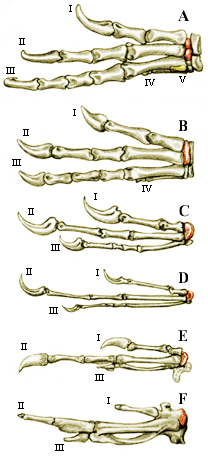
On the left is a comparison of the left manus of a variety of theropod dinosaurs all drawn so that digit II is of the same length.
The semilunate carpal and its homologues are shaded red.
A.Herrerasaurus (based on Sereno, 1993). Note that all five digits are present, which of course is primitive for the Amniota. Digit V is shaded yellow and would be hidden on the other side of the manus from this view.
B.Coelophysis (based on Colbert, 1989). Note that digit V is gone.
C.Deinonychus (based on Ostrom, 1969). Note loss of both digits IV and IV - the tetanurine condition.
D.Archaeopteryx (based on Heilmann, 1927). Note very close correspondence in proportions and relative lengths of bones to Deinonychus.
E. Hoatzin embryo Opisthocomus (based on Heilmann, 1927). Note reduction of number of bones in digit III.
F. Hoatzin adult Opisthocomus (based on Heilmann, 1927). Most of the bones of the manus fuse up in the adult, including the emilunate carpal to the metacarpals.
References:
Colbert, E. H., 1989, The Triassic dinosaur Coelophysis. Museum of Norther Arizona Bull. 57, p. 1-160.
Heilman, G., 1927, The Origin of Birds. D. Appleton and Company, 21 pp.
Ostrom, J. H.., 1969, Osteology of Deinonychus antirrhopus, and unusual theropod from the Lower Cretaceous of Montana. Peabody Musuem of Natural Histroy, Bull. 30, p. 1-165.
Paul, G. S., 1987, The science and art of restoring the life appearance of dinosaurs and their relatives:a rigorous how-to-guide. in Czerkas, S. J. and Olseon, E. C. (eds.) Dinosaurs Past and Present, Volume II, p. 6-49.
Paul, G. S., 1988, Predatory Dinosaurs of the World. Simon and Schuster, New York, 464 pp.
Sereno, P. C., 1993, The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigulastensis. Jour. Vert. Paleo., v. 13, no. 4, p. 425-450.
Is ID de wetenschappelijke (sic) uitleg voor creationiisten 150jaar Darwinisme... De evolutietheorie gewikt en gewogen David sorensen en revolutietheorie lalala DE VERRIJZENIS VAN DE JONGE VERDRONKEN KOE » Reageer (21) ID TOVERDOOS OVER SLECHTE ID-EETJES ,RESISTENTIE & TOVERDOCTORS c) OEC d) YEC
Een oeroud spoor Denisova : ZUID SIBERIË DENISOVA - mens
DMANISI AAP OF MENS DE OUDSTE ? » Reageer (1)Nakalipithecus nakayamai (<) Alweer eentje ? ..... De tand des tijds / Heidelberg-mens DE HEUPEN VAN EVA CASABLANCA MAN /erectus Hobbit is aparte soort ? » Reageer (2) NOG EEN BENDE BIJTERS POLONAISE met neanderthaler TANDEN UIT DE QESEM GROT Neanderthaler genoom Mitochondriale Genenkaart van Neanderthaler Xuchang mens
b)Biologie 1.-(EVOLUTIE ) NAS / IM Document 2008 (2) NAS / IM Document 2008 ( 1)
Cambrium & precambrium AVALON GABONESE chips
Evolutie in actie Opmerkelijk snelle adaptaties bij kroatische ruine-hagedissen:
EEN PLUIM VOOR CHINA Geef eens een pootje Pluimgewicht KLEIN DUIMPJE & DE REUS &VEREN Opnieuw gevederde Maniraptor Similicaudipteryx. Aerosteon riocoloradensis
Krokodillen
PAKASUCHUS KAPILIMAI Prestosuchus chiniquensis Terug naar zee viseter
Eritherium azzouzorum LJOEBA Darwinius masillae VLIEGENDE KATTEN ? Vleermuizen ALWEER EEN BELANGRIJK STUK UIT DE LEGPUZZEL Zee-zoogdieren ; Van de wal in de visgronden Indohyus / walvisevolutie Zeehonden-evolutie
*Blauwe en groene ( =nederlandse)teksten zijn meestal aanklikbare links *Engelse Wikipedia teksten verwijzen in de linkerkolom naar verschillende niet-engelse versies van het wikiartikel *Blauwe teksten tusssen "" , zijn voornamelijk ( gedeeltelijke)citaten afkomstig van mensen met andere meningen
Evolutie / Charles Darwin aan de basis: SELECTIE NATUURLIJK DE LEVENSBOOM UITSTERVEN DEEP TIME Biogeografie SEKSUELE SELECTIE Al jaren lang samen CO-EVOLUTIE De economie van de natuur Geleidelijke veranderingen
Unintelligent design:
IP Theorie Ons gebit , De appendix, Geboorte, De weg van het zaad , Nervus Laryngeus Recurrens, DNA , 75 mistakes , Het boek
Unintelligent design (1): Rechtop lopen Tomaso over het oog , oogspieren , Unintelligent Design (5): ZWEETVOETEN Unintelligent design (6): Vitamine C Unintelligent design (8): Kuitspieren
 Unintelligent design (1): Rechtop lopen
Unintelligent design (1): Rechtop lopen
















 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}