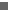

Die verscheen in het wetenschappelijk tijdschrift Science.
Aan de analyse is vijf jaar gewerkt.

Vindija cave (Croatia) waar onderstaande botfragmenten werden gevonden

Het DNA is geïsoleerd uit botten van drie Neanderthaler-vrouwen(1) ( zie foto boven ) die gevonden zijn in een grot in Kroatië.
(PZ )" ....in addition, some partial sequences were taken from other specimens, including the original type specimen found in the Neander Valley in 1856...."  PZ MYERS
PZ MYERS
(Rene fransen )
De onderzoeksgroep van Svante Pääbo en collegas heeft van drie individuen (lees: botten) het DNA geïsoleerd en geanalyseerd.
Dat is vervolgens vergeleken met DNA van 5 moderne mensen, twee uit Afrika, en een uit Europa, China en Papua Nieuw Guinea.
Waarom is het niet vergeleken met het DNA van Sapiens-tijdgenoten?
Dan zou je toch nog beter kunnen zien hoe ver ze van elkaar af stonden, en hoe gevarieerd het menselijk genoom toen was. ;
Om te bekijken of Neanderthaler werkelijk bij de Sapiens-groep behoort of een aparte tak is binnen de hominiden. ..
.
Maar Rene fransen vermoed dat
" dat veel complexer was dan vergelijken met een modern genoom. Bovendien: als er 40.000 jaar geleden onderlingen paringen waren, vind je dat eenvoudiger terug in moderne genomen, omdat de Neanderthaler genen dan de tijd hebben gekregen zich te verspreiden...." Uiteraard zou een vergelijking met tijdgenoten-genomen "op zich interessant zijn"
Een Neanderthaler-familie zoals die er mogelijk hebben uitgezien volgens de samenstellers van een expositie in het nieuwe Neanderthaler-museum in Kroatië. Foto Reuters![]()
http://tsjok45.multiply.com/photos/album/1164/Homo_neanderthalensis_
Voor de analyse van het zeer gefragmenteerde en zwaar vervuilde DNA moesten nieuwe laboratoriumtechnieken ontwikkeld worden.
Daarna kon 60 procent van het Neanderthalergenoom ontcijferd worden.
Het project stond onder leiding van Svante Pääbo van het Max Planck Instituut voor Evolutionaire Antropologie in Leipzig.
(PZ) " .....The DNA was extracted from a few bone fragments, and it was grossly degraded: the average size of a piece of DNA was less than 200 base pairs, much of that was chemically degraded, and 95-99% of the DNA extracted was from bacteria, not Neandertal. An immense amount of work was required to filter noise from the signal, to reconstruct and reassemble, and to avoid contamination from modern human DNA. These poor Neandertals had died, had rotted thoroughly, and the bacteria had worked their way into almost every crevice of the bone to chew up the remains. All that was left were a few dead cells in isolated lacunae of the bone; their DNA had been chopped up by their own enzymes, and death and chemistry had come to slowly break them down further..." .PZ MYERS
De Neanderthaler (Homo neanderthalensis) is de naaste verwant van de moderne mens Homo sapiens.
Hij leefde tot 30.000 jaar geleden, van West-Europa tot Zuid-Siberië.
Een analyse van het Neanderthaler-DNA biedt de beste mogelijkheid om te bepalen welke genetische aanpassingen typisch zijn voor de moderne mens.
Daarnaast verheldert het onderzoek de geschiedenis van de Neanderthaler en de moderne mens.
Homo sapiens heeft zijn voorganger niet als een Blitzkrieg verdrongen, want de twee soorten mengden immers.
In een eerste analyse ( van het mitochondriale mtDNA )die Pääbo in 2006 publiceerde, was zon genetische bijdrage juist niet gebleken (1) .


Richard E. (Ed) Green, Svante Pääbo met Neanderthaler schedel
( Rene fransen ) :
" .....Maar de analyse van genoom-DNA sluit toch aan bij eerdere analyses van mitochondriaal DNA, in de zin dat Neanderthalers duidelijk verschillen van moderne mensen.
Er is overeenkomst, maar dat valt te verwachten gezien de recente splitsing.
Een caveat is, dat het nog om kleine aantallen gaat.
En aangezien op een willekeurige plek in het DNA de verschillen tussen twee moderne mensen soms groter zijn dan tussen een moderne mens en een Neanderthaler, maakt dat de conclusies voorlopig.
Al laten de onderzoekers door verschillende vergelijkingen naar mijn idee wel overtuigend zien, dat de Neanderthaler geen homo sapiens was.
De Leidse Neanderthaler-specialist prof. Wil Roebroeks zegt:
Zeker in de Verenigde Staten was de consensus onder deskundigen: er was geen bijmenging.
Dat blijkt nu een misvatting.
Vooral niet-Afrikanen (Europeanen, maar ook Chinezen en Papoeas) dragen de sporen bij zich van prehistorisch contact met Neanderthalers. 1 tot 4 procent van het DNA van de niet-Afrikanen is afkomstig van Neanderthalers.

http://www.sciencemag.org/cgi/content/full/328/5979/710 (Abstract )
"....Neandertals, the closest evolutionary relatives of present-day humans, lived in large parts of Europe and western Asia before disappearing 30,000 years ago. We present a draft sequence of the Neandertal genome composed of more than 4 billion nucleotides from three individuals. Comparisons of the Neandertal genome to the genomes of five present-day humans from different parts of the world identify a number of genomic regions that may have been affected by positive selection in ancestral modern humans, including genes involved in metabolism and in cognitive and skeletal development. We show that Neandertals shared more genetic variants with present-day humans in Eurasia than with present-day humans in sub-Saharan Africa, suggesting that gene flow from Neandertals into the ancestors of non-Africans occurred before the divergence of Eurasian groups from each other...."
( Rene Fransen )
".....De bottomline is, dat er aanwijzingen zijn dat er zeer sporadisch sprake is geweest van uitwisseling van genen (dus: er is onderling gepaard). Zon 1-4 procent van het DNA van de niet-Afrikaanse proefpersonen in de studie kan afkomstig zijn van Neanderthalers. Voor de Afrikanen geldt dat niet. De meest aannemelijke verklaring, aldus de onderzoekers, is dat de moderne mens bij emigratie uit Afrika Neanderthalers tegenkwam (logisch gezien in het Midden Oosten) en vervolgens bij verdere migratie wat genen heeft meegenomen.
Het contact tussen" neanderthaler" en de uit afrika emigrerende "archaische " homo sapiens bestond, volgens de onderzoekers, in het Midden-Oosten, in de periode tussen 80.000 ( a 100.000 ) en 50.000 jaar geleden.
Roebroeks:
Dat klopt met archeologische vondsten in Israël. Daar zie je vanaf 100.000 jaar geleden eerst moderne mensen, en daarna Neanderthalers. En ze deden hetzelfde: ze gebruikten dezelfde stenen werktuigen.
Dat is al heel lang een puzzel.
De onderzoekers identificeren bovendien twintig DNA-fragmenten, met daarop een vijftigtal genen, die belangrijk zijn geweest bij de ontwikkeling van de moderne mens.
( Rene Fransen ) ."....Er een flink aantal plekken in het DNA die uniek zijn voor de moderne mens, en die volgens de onderzoekers dus zijn ontstaan na de splitsing van Neanderthaler en moderne mens, geschat op 400.000 jaar geleden.
Een aantal mutaties die uniek zijn voor moderne mensen zijn gevonden in genen die een rol spelen bij de cognitieve ontwikkeling (denkvermogen, hersenen) en skeletvorming (ribben, sleutelbeen).
Die laatste verschillen zijn niet verrassend, want dat de skeletten verschillen wisten we natuurlijk al. "

![]()
![]()
![]()
http://tsjok45.multiply.com/photos/album/1164/Homo_neanderthalensis_
300.000 à 400.000 jaar geleden zijn de voorouders van de mens afgesplitst van de voorouderlijke "common ancestor" -tak van de Neanderthalers, legt Ronny Decorte van het departement Menselijke Erfelijkheid aan de KU Leuven uit.
De Neanderthalers vertrokken richting Midden-Oosten en Europa en zijn daar gebleven tot ze 30.000 jaar geleden om een of andere reden verdwenen zijn.
De homo sapiens - de mens, dus - ontwikkelde zich ondertussen 150.000 à 200.000 jaar geleden in oostelijk Afrika.
Dat was een kleine groep van zon 2.000 tot 10.000 mensen. Het waren jagers en verzamelaars en ze moesten dan ook migreren om aan voedsel te geraken, aldus Decorte.
Zowat 70.000 jaar geleden zijn ze vanuit Afrika in het Midden-Oosten beland. (2)
Daar woonden op dat moment Neanderthalers, en blijkbaar hebben beide groepen sexueel contact gehad .
(Rene fransen ) Waarom is er maar beperkt gepaard tussen de twee? Dat kan tal van redenen hebben.Of het toevallig is, is lastig te zeggen. Want waar zet je het tegen af?
Dat verklaart meteen waarom er bij Afrikaanse groepen geen invloed is, stelt Decorte.
De vermenging is wel al heel vroeg gebeurd, vooraleer de groep zich opsplitste in een Aziatisch en een Europees gedeelte, anders kon er bij de Aziatische volkeren geen spoor meer worden teruggevonden. De hoeveelheid DNA doet vermoeden dat het om een heel beperkt contact ging.
Grotere herseninhoud
Een onenightstand met 4 procent Neanderthaler-DNA in onze genen tot gevolg dus.
En die vaststelling mag dan al historisch en evolutionair gezien belangrijk en hoogst opmerkelijk zijn, ze zal waarschijnlijk geen wezenlijke gevolgen hebben.
Daarvoor is het percentage te klein, zegt Decorte.
Bovendien verschilden de Neanderthalers uiterlijk wel van de mens, qua gedrag was er vermoedelijk niet zo heel veel verschil. De Neanderthalers hadden een grotere herseninhoud dan de mens en ze hadden ook al een eigen cultuur.
Zo werden de overledenen begraven, net zoals bij de mens. (3) 
Toch sluit het Duitse onderzoeksteam niet uit dat er wel degelijk een impact is.
Een belangrijke volgende stap is nagaan of de genen van de Neanderthalers een betekenis hebben. Met andere woorden: is er een biologisch verschil tussen Afrikanen en de rest van de wereld, dat verklaard kan worden door het Neanderthaler-DNA?,
aldus de onderzoekers.
Bronnen en links
Rene Fransen http://www.sterrenstof.info/?p=1040
kennislink http://www.kennislink.nl/publicaties/genoom-neanderthaler-ontrafeld
Carl Zimmer
http://blogs.discovermagazine.com/loom/2010/05/06/skull-caps-and-genomes/
Jerry Coyne :
http://whyevolutionistrue.wordpress.com/2010/05/06/we-carry-neanderthal-genes/
http://whyevolutionistrue.wordpress.com/2010/05/07/quick-guide-to-the-neandertal-genome/
Er is trouwens nog een mogelijk "derde " tak naast de Neanderthaler , de sapiens ---> De Homo uit "Denisova "
http://whyevolutionistrue.wordpress.com/2010/05/13/o-noes-another-species-of-human-in-eurasia/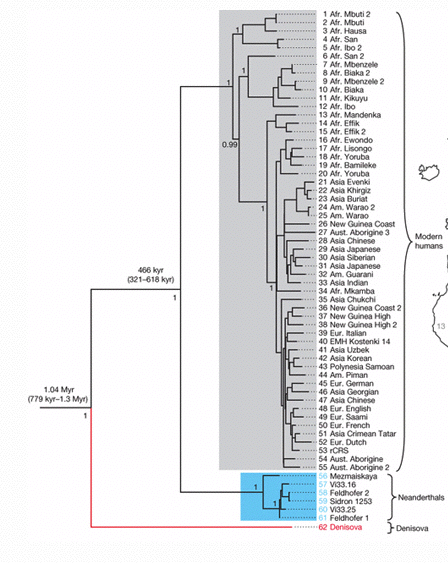
PZ Meyers 
http://pandasthumb.org/archives/2010/05/neandertal.html
http://scienceblogs.com/pharyngula/2010/05/neandertal.phpPZ MYERS
zie ook <-- ( klik) Denisova : ZUID SIBERIË<-- (klik) Mitochondriale Genenkaart van Neanderthaler
John Hawks
http://johnhawks.net/weblog/reviews/neandertals/neandertal_dna/neandertals-live-genome-sequencing-2010.html
http://www.nytimes.com/2010/05/07/science/07neanderthal.html Creationisten leuter
Creationisten leuter
1.-
POLONAISE met neanderthaler
Creationisten benadrukken zoals kon worden verwacht :
( Rene fransen ) ." ...de overeenkomsten en het feit dat er wel degelijk onderlinge voortplanting lijkt te zijn geweest.
Maar ..... dat was op beperkte schaal, en het past in het beeld van twee ondersoorten die een gemeenschappelijke voorouder delen dat er (nog) geen volledige seksuele isolatie is opgetreden.
Want nauw verwant zijn ze natuurlijk."
2- http://www.vkblog.nl/bericht/314204/Hot_news%21_Ook_Svante_P%E4%E4bo_moet_eraan_geloven...
PeeBee :
" ....Dacht men voorheen altijd dat de twee zich apart hadden "entwickelt", het is nu duidelijk dat we wel degelijk( 1.)- met maar een soort te maken hebben. ....
(2.- )Neandertalers en Homo sapiens hebben gewoon met elkaar gepaard en nakomelingen gekregen ...
Als twee organismen kunnen paren en samen nageslacht kunnen voortbrengen, behoren ze volgens de species-definitie van Ernst Mayr tot een en dezelfde soort.
(3)Leek het mtDNA nog aanwijzingen te bevatten dat de twee mensentypen niet tot dezelfde soort zouden behoren, de( nu uitgevoerde ) genomische analyses zijn eenduidig.
Eigenlijk zijn uitgestorven Neandertalers en alle hedendaagse moderne mensen gewoon de derivaten van een en hetzelfde baranoom."
Het doet er niet toe voor het al dan niet kloppen van de (moderne) evolutietheorie
Het maakt voor de evolutie theorie niet uit of Sap en neanderthaler van dezelfde soort waren of niet.
Om weer een vergelijking te maken met zwaartekracht, als het nu na al die jaren blijkt dat g niet 9,81 maar 9,82 is op aarde, dan zegt dat niets over de zwaartekracht theorie
.
Het rare is, dat ik ook niet zie hoe het iets over peebee's GUToB( of andere creationistische "theorieen " ) zal zeggen. Misschien dat ik het gemist heb, maar wat zorgt ervoor dat voorspeld wordt dat neandertalis dezelfde soort is?
Flut-wetenschap
( Rene) .....de basisgegevens van de studie van Pääbo passen sowieso niet binnen de spelregels van de leefwereld van (YEC creationisten , inzonderheid ) PB.
Pääbo stelt dat er "some interbreeding" tussen Homo sapiens en (proto) Neanderthalers plaatsvond tussen 80.000 en 50.000 jaar geleden. Maar toen bestond de aarde volgens PB niet eens!
PB( en YEC's )zullen dus terug moeten naar het basismateriaal, om het binnen hun eigen "paradigma" te duiden (waaronder dateren), en op grond van de zo verkregen gegevens een argumentatie moeten opbouwen waarmee ze hun eigen "theorie" kunnen onderbouwen, of de - volgens hen enige - andere theorie falsifiëren.
Zo kan (bijvoorbeeld ) PB op grond van de opgewerkte gegevens van Pääbo's studie helemaal niet concluderen dat "daarmee dan een creationistische stelling wordt bevestigd en een evolutionistische stelling verworpen".
Binnen het naturalistische "paradigma" is er
ten eerste
geen "evolutionistische" (hé, da's een nieuw woord) stelling verworpen,
en ten tweede
zal PB eerst opnieuw data moeten gaan interpreteren binnen zijn eigen genesiaanse "paradigma" voordat hij ook maar een begin gemaakt heeft van een creationistisch verificatie-bewijs. (En dat laatste is argumentatief al erg zwak.)
Heeft iemand PB wel eens verteld dat hij een flutwetenschapper is die niet eens logisch kan redeneren?
Hij is inderdaad een schoolvoorbeeld van (om het in zijn eigen woorden te zeggen :) " een Schetenwapper "
1.-
Men heeft goede redenen om te (veronder )stellen dat : De Neanderthaler helemaal niet wereldwijd verspreid voorkwam ....
Uiteraard niet in de de Amerika's ( waar alleen oude prehistorische aziatische " homo sap" vertegenwoordigers als eersten ( geologisch ) vrij recent arriveerden )
De Neanderthaler is ( tot vandaag ) ook niet gevonden in "sub-sahara Afrika"( = vroeger : "zwart" afrika )
In zuid oost afrika ( een woonplaats ( minstens 70.000 jaar oud ) van de vroege homo sap ?--> plombos grot ) ontbreekt hij volledig
Ook in de hoorn van Afrika / Ethiopië ( zie hieronder ) Eritrea ( minstens 125.000 jaar oud ) , en wijde omgeving (De bakermat van de sapiens ?) is tijdgenoot Neanderthaler ( nog ) niet gevonden , de oude homo sapiens dus wél )
Vroege Neanderthalers zaten wel in ( sporadisch ) noord -afrika en waren goed gesettled in het Midden Oosten
http://evodisku.multiply.com/journal/item/688/Homo_sapiens
![]()
out of africa /Multiregional theorie

 |
|
In 2003 werden in de Midden Awash-vallei in Ethiopië drie Homo Sapiens schedels gevonden van 160.000 jaar oud. Het zijn de oudste goed gedateerde schedels van de anatomisch moderne mens die tot nu toe zijn gevonden.

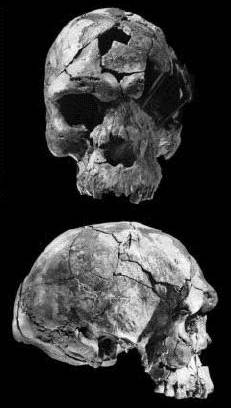
foto links / 2001 David L. Brill Brill Atlanta
FOSSIL: Homo sapiens idaltu, BOU-VP-16/1
VIEW: Frontal
SITE & LOCATION: Upper Herto Member, Bouri peninsula, Middle Awash, Ethiopia
FOSSIL CREDIT: Housed in National Museum of Ethiopia, Addis Ababa
http://evodisku.multiply.com/journal/item/500/Fossiele_hominiden
* Er zijn in totaal ongeveer +400 verschillende & belangrijke fossiele neanderthaler individuen gevonden
zie -->
|
|
-De echte antropologische wetenschap ( niet de populaire "boekskes " ) beweerde al lang =
"Neanderthals are either classified as a subspecies of humans (Homo sapiens neanderthalensis) or as a separate species (Homo neanderthalensis).[3] "
ref [3]: Tattersall I, Schwartz JH (June 1999). "Hominids and hybrids: the place of Neanderthals in human evolution". Proceedings of the National Academy of Sciences 96 (13): 71179.
Alleen maar omdat ze konden paren betekent niet dat ze van dezelfde soort waren. Als soort A met soort B kan paren, en soort B met soort C, kan het voorkomen dat soort C niet kan paren met soort A (en.wikipedia.org/wiki/Ring_spe...). Ze hoeven dus niet per se van dezelfde soort te zijn als ze met elkaar kunnen(of niet kunnen ) paren.met fertiel nageslacht als resultaat
Bovendien zijn er gedocumenteerde gevallen van (hypothetische ) soortvorming die juist door soort -bastaardering zijn onstaan
http://www.ucl.ac.uk/taxome/hyb/
Waaruit deze aanhef als teaser :
"One of the best ways of showing evolutionary continuity between species and geographic races is to demonstrate that hybridization still occurs between closely related species. In Victorian times and early this century, naturalists were very interested, like "stamp-collectors", in freaks of nature, including rare hybrids between species. Between the 1930s and about 1980, there was decreased interest about the peculiarities of nature, and increased emphasis on the "fundamental" biological "realities" of animal species. (Hybridization between plant species is so abundant and easily shown, of course, that this rather myopic view of "pure", "good" species has never really caught on among botanists). Widely used field guides from this period often omit pictures even of common hybrids of birds and butterflies that can be seen in the wild, while treating much rarer or even extinct species in the same book. Recently, however, there has been renewed interest in all aspects of biodiversity, including that within species, and it is possible to discern a return of interest in variants, hybrids, and exceptions ("bad species") as well as the good species. In some beautiful recent books, often thorough world treatments of particular groups of butterflies and birds, hybridization between species is again becoming well-documented. ..." (Lees verder op de pagina )
Natuurlijk iets anders dan = allemaal afkomstig van hetzelfde "geschapen" baranoom
2.-
Ja ... en ?
De biologische soortbegrip definitie van Mayer is een biologische heuristiek : het is één van de vuistregels gebruikt door taxonomen : het is zeker geen altijd geldende wet ... zoals steeds in de biologie zijn er veel uitzonderingen
PB : " De Neandertaler kan volgens de modernste genetische analyses niets anders zijn dan Homo sapiens."
( Roel ) Dit is al te kort door de bocht.
-Ze paarden met onze voorouders; dat is een heel andere conclusie.
-Trouwens, afhankelijk van de definitie, zou zelfs een krop sla tot de Homo sapiens kunnen behoren...
Veel geblaat en weinig wol weer.
3.- de mitochondriale verschillen tusszen "homo sap "en "neanderthaler " zijn niet plots verdwenen .... het betekent misschien ( volgens mij ) o.a. nog altijd dat de neanderthaler en de homo sap zeker niet dezelfde mitochondriale Eva delen :
Alle homo sap doen dat echter wél ( ook de San en de afrikanen .....die echter niets van doen hebben met neanderthaler )(1)
http://en.wikipedia.org/wiki/Mitochondrial_Eve
http://nl.wikipedia.org/wiki/Mitochondriale_Eva
Over minder efficiente neanderthaler mitochondrieen en ( erbij horende ?) astenozoospermie
http://weetlogs.scilogs.be/index.php?op=Default&Date=201001&blogId=8
 Had de neanderthaler minder bewegelijk sperma ?
Had de neanderthaler minder bewegelijk sperma ? 
http://tsjok45.multiply.com/photos/album/1164/Homo_neanderthalensis
3.-
de eerlijkheid gebied om niet alle creationisten over één kam te scheren _
zo heb je hier bijvoorbeeld wat creationist Todd Wood (een echte "baraminoloog " !!) ervan vindt:
toddcwood.blogspot.com/2010/05...
<-- (klik) samenvatting van het " Green & al paper" Creationist Todd Wood vindt van de Neanderthaler : het is geen homo sapiens
(Todd Wood is de creationist die een poos geleden riep: er is wel degelijk bewijs voor evolutie - maar toch blijf ik creationist).
NOTEN
(1)
<-- (klik) Mitochondriale Genenkaart van Neanderthaler Noorderlicht ;
"....Het mitochondriale DNA van de neanderthaler verschilt zodanig van dat van moderne mensen, dat het ver buiten de variatie binnen onze eigen homo sap soort valt.
Alle mensen die nu leven, hebben hun mitochondriën geërfd van een vrouw die omstreeks 150 duizend jaar geleden leefde.
De laatste vrouw waarvan zowel wij als de neanderthaler mitochondriën hebben gekregen, zal ongeveer 660 duizend jaar geleden hebben rondgelopen, met een foutenmarge van 140 duizend jaar. Daarmee kan het idee dat de moderne mens een scheutje neanderthalerbloed heeft, naar de prullenbak. Tenzij de vermenging alleen van vaderskant heeft plaatsgevonden, want dan is dat niet te zien in het mitochondriaal DNA.
Wel in de rest van het erfelijk materiaal ( de nucleus ) ..."
- Dit (oorspronkelijke ) onderzoek was gebaseerd op één bot van één enkel (kroatisch ) individu
- zijn er ook nog andere mtDNA onderzoekresultaten van de botten van de drie kroatische vrouwelijke neanderthaler botten ?
- Het blijft zo dat er regionale verschillen ( = rassen dus ) zijn binnen de Neanderthalers van west naar oost...maar die zijn volgens vervolgonderzoek naar het mtDNA (in 2008 )veel kleiner dan vroeger was verwacht
op basis van fragmenten van mitochondriaal neanderthaler mtDNA
Weinig genetische variatie tussen Neanderthalers :NRC 10 augustus 2008 (Door Hendrik Spiering)
"..... De genetische variatie bij Neanderthalers was veel kleiner dan bij moderne mensen (die ook al niet groot is).
Dat is de belangrijkste conclusie die een team van genetici trekt uit
de reconstructie van het complete mitochondriaal genoom van de Neanderthaler (Cell, 8 augustus) .
Deze reconstructie van het genetisch materiaal uit de mitochondriën (mt-DNA) van een 38.000 jaar oude Neanderthaler uit een grot in Kroatië is op zichzelf al een grote stap vooruit, tot nu toe waren er alleen korte fragmenten van het mtDNA bekend. Het is het eerste vrijwel foutloze genoom van archaïsch DNA, zegt de trotse hoofdonderzoeker Richard Green van het Max Planck Instituut in Leipzig. Maar het mag ook gelden als generale repetitie voor de veel complexere reconstructie van het complete kern-DNA van de Neanderthaler.
( waarvan dus nu ongeveer 60 % is bekend in een voorlopig referentie- model gebaseerd op drie fossiele individuen )
Mitochondriën zorgen voor de energievoorziening in de cel en omdat ze miljarden jaren geleden zijn ontstaan als ingesloten bacteriën, bezitten ze nog altijd een miniem eigen genoompje. De Neanderthaler had 16.565 baseparen in zijn mtDNA, vier minder dan de moderne mens. Zijn kern-DNA zal ongeveer 3 miljard baseparen beslaan, evenveel als de moderne mens. In de eerste analyses van dat kern-DNA bleek al dat het Neanderthaler-DNA voor 99,5 à 99,9 procent overeenkomt met het moderne DNA. De ongeveer 25.000 jaar geleden uitgestorven Neanderthaler is de meest naaste verwant van de moderne mens.
De geringe genetische variatie van de Neanderthaler leiden de onderzoekers af uit het type mutaties in dertien genen die het mtDNA telt. De Neanderthaler heeft daarin relatief veel mutaties die invloed hebben op het resulterende eiwit. Dat hangt meestal samen met een kleine bevolkingsomvang.
In een kleine bevolking heeft natuurlijke selectie weinig invloed op het genoom en worden negatieve mutaties (en de meeste eiwitveranderende mutaties zijn negatief) worden niet uitgewied.
En een kleine bevolking( grotere kans op inteelt ? ) hangt altijd samen met een geringe genetische variatie.
Ongetwijfeld hangt de geringe bevolkingsomvang samen met de harde omstandigheden in het leefgebied van de Neanderthalers: Europa en West-Azië tijdens de IJstijden.
De geschatte bevolkingsomvang van de Neanderthalers lag overigens al rond de 3 à 4.000.
Verder heeft de moderne mens in totaal meer mutaties in zijn mtDNA dan de Neanderthaler (75 tegen 64, maar daarvan zijn minder niet-eiwitveranderende dan bij de Neanderthaler: 57 tegen 44). De reden voor dat grotere aantal is onbekend.
Verder valt uit vergelijking met het menselijk mtDNA te concluderen dat de laatste gemeenschappelijke moeder van mensen en Neanderthalers ergens rond 600.000 jaar geleden moet hebben geleefd mtDNA erft alleen via de vrouwelijke lijn over. Dat tijdstip komt redelijk overeen met andere schattingen, die allemaal zo rond het half miljoen jaar liggen. Uit de toen levende mensachtige Homo heidelbergensis ontstond in Europa de Neanderthaler, in Afrika Homo sapiens. "
De tand des tijds / Heidelberg-mens
P.S.
als je ver genoeg terug gaat in de tijd ( tot op hun common ancestor uiteraard ) zijn sap en "neanderthaler "natuurlijk wél van dezelfde soort ... maar ze bezitten in elk geval niet dezelfde mitochondriale eva ...
.Euh .... Neanderthaler verdronk in de zondvloed ?
uit het onderzoek-artikel :
"....Although gene flow from Neandertals into modern humans when they first left sub-Saharan Africa seems to be the most parsimonious model compatible with the current data, other scenarios are also possible.
For example, we cannot currently rule out a scenario in which the ancestral population of present-day non-Africans was more closely related to Neandertals than the ancestral population of present-day Africans due to ancient substructure within Africa (Fig. 6).
If after the divergence of Neandertals there was incomplete genetic homogenization between what were to become the ancestors of non-Africans and Africans, present-day non-Africans would be more closely related to Neandertals than are Africans.
In fact, old population substructure in Africa has been suggested based on genetic (81) as well as paleontological data (86)...."
(Gerdien De Jong )
Uit het onderzoek blijkt, als ik het wel begrepen heb, dat de Neanderthaler een paar honderd duizend jaar geleden nog soortgenoot van de Homo Sapiens was.
Een paar honderdduizend jaar geleden hadden de Neanderthaler en de anatomisch moderne mens een gemeenschappelijke voorouder.
Homo sapiens is niet erg oud, vermoedelijk minder oud dan de Neanderthaler. Verder hangt hier veel af van de persoonlijke voorkeur voor te hanteren soortsbegrip - soort is biologisch niet altijd scherp, en er bestaan vele soortsbegrippen
(2) Een" andere " groep van de oost-afrikaanse homo sapiens , emigreerden naar zuidelijk afrika ?
(3) ![]()
![]()
![]()
![]()
![]()
http://tsjok45.multiply.com/photos/album/1164/Homo_neanderthalensis_
De Neanderthalers bezaten daadwerkelijk een cultuur : er zijn sieraden ontdekt :
in Vogelherd ( en Slovakije ) zijn benen fluiten ,( waarschijnlijk) symbolische sculpturen (vruchtbaarheidssymbolen ) en andere artefakten gevonden : maar inhoeverre de europese vondsten , produkten zijn van een echte mengcultuur (of produkten van wederzijdse imitatie ) is nog steeds niet bekend ... feit is dat de laatste Neanderthalers sporen van een wel merkwaardige "culturele versnelling " hebben nagelaten zo'n 40.000 jaar geleden begonnen ( Het tijdperk dat de homo sap zich reeds stevig had gevestigd in Europa ) ![]()
http://tsjok45.multiply.com/photos/album/234/Homo_sapiens_in_europa_archeo-antropo
http://evodisku.multiply.com/journal/item/682/Homo_sapiens_in_europa

Divje Babe flute ( slovakije)
VOgelherd
Oudst bekende Venus 35.000 jaar oud Hohle Fels-grot, in de Zwabische Jura. /
Waarschijnlijk homo sapiens
-Dat de neanderthalers hun doden begroeven en van grafgiften ( = bloemen ) voorzagen is voornamelijk gebaseerd op vondsten in Schanidar (irak )
Maar nader onderzoek van het kindgraf in Noord-Irak dat hiertoe als inspiratie diende, leverde op dat het verzamelde stuifmeel in het graf (de restant van de grafbloemen), ook veroorzaakt kan zijn door de wind of door overijverige woestijnratten,
Een mooi verhaal moet je niet kapot checken, natuurlijk .....
Toch blijken andere vondsten( althans er wordt steeds gewag van gemaakt maar dat wordt naar mijn weten bijna nooit nader gespecifeerd ) de voormelde stelling te ondersteunen
http://www.kunstinderegio.nl/upload/Expo/Teksten%20begeleidend%20boekje%20tt%20Neanderthalers.pdf
- De gevonden Neanderthaler sequenties kunnen de studie van de menselijke evolutie verder ophelderen
" Gelijkaardige genetische sequenties die voorkomen bij chimp , orang oetang en Neanderthaler, maar die verschillen van de menselijke varianten betekenen dat die homologe homo sap - sequenties , specifiek soorteigen moeten zijn "
zegt Burbano
" deze studie hielp daarom ook bij het samenstellen van een begin van een genetische catalogus met specifieke menselijke substituten cvoor die sequenties "
PZ MYERS
" ..... these are 78 genes that have changes in their coding sequence. There are also several hundred other non-coding, presumably regulatory, sequences that are unique to humans and are fixed throughout our population. To the evo-devo mind, these might actually be the more interesting changes, eventually but right now, there are some tantalizing prospects in the coding genes to look at.
Some of the genes with novel sequences in humans are DYRK1A, a gene that is present in three copies in Down syndrome individuals and is suspected of playing a role in their mental deficits; NRG3, a gene associated with schizophrenia, and CADPS2 and AUTS2, two genes associated with autism. These are exciting prospects for further study because they have alleles unique and universal to humans and not Neandertals, and also affect the functioning of the brain. However, let's not get confused about what that means for Neandertals. These are genes that, when broken or modified in modern humans, have consequences on the brain. Neandertals had these same genes, but different forms or alleles of them, which are also different from the mutant forms that cause problems in modern humans. Neandertals did not necessarily have autism, schizophrenia, or the minds of people with Down syndrome! The diseases are just indications that these genes are involved in the nervous system, and the differences in the Neandertal forms almost certainly caused much more subtle effects.
Another gene that has some provocative potential is RUNX2. That's short for Runt-related transcription factor 2, which should make all the developmental biologists sit up and pay attention. It's a transcription factor, so it's a regulator of many other genes, and it's related to Runt, a well known gene in flies that is important in segmentation. In humans, RUNX2 is a regulator of bone growth, and is a master control switch for patterning bone. In modern humans, defects in this gene lead to a syndrome called cleidocranial dysplasia, in which bones of the skull fuse late, leading to anomalies in the shape of the head, and also causes characteristic defects in the shape of the collar bones and shoulder articulations. These, again, are places where Neandertal and modern humans differ significantly in morphology (and again, Neandertals did not have cleidocranial dysplasia they had forms of the RUNX2 gene that would have contributed to the specific arrangements of their healthy, normal anatomy).
These are tantalizing hints to how human/Neandertal differences could have arisenby small changes in a few genes that would have had a fairly extensive scope of effect.
Don't view the many subtle differences between the two as each a consequence of a specific genetic change; a variation in a gene like RUNX2 can lead to coordinated, integrated changes to multiple aspects of the phenotype, in this case, affecting the shape of the skull, the chest, and the shoulders....."
De wetenschappers hebben specifieke genen gevonden die de evolutie van schedel- , sleutelbeen-en de ribbenkast-kenmerken , aansturen ...
Verandering (= mutatie ) in het RUNX2 gen bijvoorbeeld , lijkt een voortreffelijke rol te hebben gespeeld in de unieke bovenlijf en schedel opbouw van homo Sap , iets wat ten sterkste verschilt van de Neanderthaler
-Herseninhoud / meer hersencellen ?
-Overigens is een grotere herseninhoud ( en volgens de allometrie --> het EQ : http://nl.wikipedia.org/wiki/Encefalisatie-quoti%C3%ABnt ) geen indicatie van een "betere " kwaliteit van de hersen- en cultuur- produkten ... zie bijvoorbeeld Homo floresiensis
- de " gemiddelde " inhoud van de neanderthaler schedel ( op basis van 400 exemplaren ) valt trouwens binnen de "hogere" variatiewaarden van de Homo Sap schedels ( van 1000 tot 2000 cc ) ...











































 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}
{kind=link}
{kind=link}