Dit blog is een onderdeel van EVODISKU WAT IS DE BEDOELING EN WAAR STAAT DIT BLOG VOOR
****Wie meent dat alles inmiddels wel over de evolutietheorie gezegd is en dat de discussie gesloten kan worden, ziet over het hoofd dat de wetenschap niet stil staat.
***Wie meent dat inmiddels het creationisme definitief het pleit heeft verloren en dat de discussie gesloten kan worden , ziet over het hoofd dat het "creationisme" is geevolueerd ( en zal evolueren ) in nieuwere mimicrytische vormen( meme-complexen ) zoals bijvoorbeeld het ID(C)
***Dit blog is speciaal opgezet om de aktualiteit binnen het evolutie-creationisme debat te volgen en van kommentaren te voorzien ... waartoe de lezers zijn uitgenodigd bij te dragen ...
Let echter wél op het volgende :
"Je bent een rund als je hier met religie stunt " ....
24-03-2008
De zogenaamde levende fossielen
WAT IS EEN LEVEND FOSSIEL ( LF) NU EIGENLIJK ? dat is geenszins voor iedereen even duidelijk , me dunkt .....
Hoe definiëren we best en sluitend (paleontologisch /geologisch en volgens de evolutiebiologie )een 'levend fossiel'? en wat zijn de (eventuele )leken-definities , taalkundige valkuilen , oorspronkelijke naamgevingen en omgangstaal-betekenissen ,die hebben geleid tot verwarring en misvattingen .
.-Betekent "levend fossiel " gewoon dat = een extant type / soort (met of zonder extante verwanten) fossiel identieke( of vrijwel identieke) organismen met gelijkaardige vormgeving kent ?. Dat lijkt toch wel een oppervlakkige , gemakkelijke en ietwat domme want vooral overbodige , definitie ; immers meer dan 50% van de nog levende tetrapoden soorten ( om slechts maar die te noemen ) hebben ( voorspelbaar ) , vergelijkbare /(enigzins) identieke fossiele vertegenwoordigers. (1)
Een generaliserende definitie is erg leuk en zeker voldoende .... voor zolang je er niet dieper over begint na te denken.
De redenering was toen dat deze organismen al " bekend "waren ,voor hun ontdekking , vanuit het Fossielen-archief en dat die bekende fossielen ( toen ) niet (veel )verschilden ___ op het eerste zicht ___ van de gevonden levende "soorten"(?) .
Je kan dat eigenlijk evengoed beweren voor zowat alle nu levende soorten . Er bestaan fossiele Homo sapiens , Canis lupus, Corvus corvus en Giraffa camelopardalis.... enz ... Betekent dit dat ze allemaal levende fossielen zijn ?
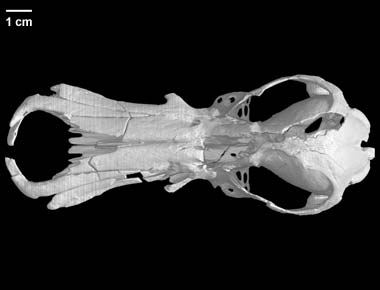
*Sphenodon punctatus is bijvoorbeeld een zogenaamd " levend fossiel " maar is toch als soortkompleet onbekend uit het fossielen-archief . Het is bovendien een extant en zeer gespecialiseerd reptiel dat slechts aan een zeer specifiek millieu optimaal is aangepast ( onwaarschijnlijk dus dat deze soort al lang "onveranderd " rondloopt )(2) ....en erg verwarrend allemaal wanneer het gaat over onze definitie-pogingen
Misschien is de term LF alleen maar geldig ' wanneer een soorten-groep( van hogere orde dan een soort of een aantal ondersoorten) als morfologisch "onveranderd " is bekend gebleven gedurende een BEPAALDE LANGERE GEOLOGISCHE TERMIJN vanaf hun eerste opduiken in het fossielen-archief (m.a.w. in het bezit van een fossiele geschiedenis die minstens terug gaat tot ( bijvoorbeeld ) voor het Pleistoceen --->[de Kassuarissen stammen uit het Plioceen, thylacines zijn Pleistoceen die waren toen tot 25% groter dan de recente (ondertussen ook uitgestorven )voorbeelden., balein walvissen verschenen voor het eerst in het Oligoceen]). ..... En gedurende dat ganse tijdperk / tot op heden ( in de overblijvende vertegenwoordiger /extante soort) moet dat morofologische bauplan 'ongewijzigd' blijven ....
* Platypi zijn dan volgens die betekenissen/criteria , uitgesloten: omdat er fossiele specimen bestaan uit die groep die heel anders zijn dan de moderne Ornithorhynchus anatinus zoals : Teinolophos
Science 23 November 2007: Vol. 318. no. 5854, p. 1237 News Focus SOCIETY OF VERTEBRATE PALEONTOLOGY MEETING: Jaw Shows Platypus Goes Way Back
......Het fossiel archief is erg spaarzaam wat betreft de Monotremata ( = Monotremen = Cloacadieren) Het oudst bekende specimen isbekend van één enkele tand ,ongeveer -62 Ma en gevonden in Patagonië :de tand heeft de typische gedrongen vorm eigen aan platypii ( = Vogelbekdieren )voordat hun stamlijnen voorgoed hun tanden verliezen .
Een nieuwe analyse van fossiele kaken gevonden in Australië stelt dat de vondst afkomstig is van een platypus die minsten - 112 ma geleden leefde. "Dat is zeer oud,voor een monotremen- soort " zegt Timothy Rowe van de Universiteit van Texas (UT), Austin,.
Teinolophos trusleri werd ontdekt in 1997 dichtbij Inverloch, Australië, en werd beschreven door Thomas Rich van het Victoria Museum Victoria (Melbourne),Pat Vickers-Rich van Monash University , en collega's. Het specimen bestaat uit een (onder)kaken en tanden.
"Grand canal. " Het grijze gedeelte op de CT scan ( boven ) van de kaak , markeert de zenuwbanen
Op zoeken naar meer anatomische aanwijzingen over de evolutie van deze zoogdieren, liet het team van Rich de fossiele kaken onderzoeken door Rowe, (een paleontoloog die toegang heeft tot de diensten van een team die beschikken over scan -toestellen van de UT Austin ). Het aftasten van de drie specimen leverde het beeld op van een groot inwendig zenuw-kanaal( groeven in het bot ) over de volledige lengte van de kaak,; Net zoals het zenuw-kanaal( = in een moderne platypus die de zenuwvezels van de electrosensorische zintuigelijke uit de gevoelige bek verbindt met de hersenen "Geen enkel ander zoogdier bezit een dergelijk zenuwkanaal van dezelfde omvang ," vertelde Rowe nog . Zelfs in het vroege Krijt lijken platypii reeds electrosensorische zintuigen in de "bek" te hebben gebruikt . "Dit is de( tot nu toe )meest dwingende aanwijzing ( =evidentie ) voor de stelling dat Teinolophos behoort tot de familie der vogelbekdieren ."
Nota =De oudere vogelbekdierachtigen, Steropodon, Kollikodon en Teinolophos, behoren mogelijk ook tot de Familie Ornithorhynchidae (Vogelbekdieren) , maar worden er regelmatig buiten geplaatst.
links schedel van Obdurodon dicksoni,rechts schedel van moderne Ornithorhynchus anatinus
Een significant morfogenetisch/anatomisch verschilpunt tussen moderne en fossiele vormen in de stamlijnen van deze dieren is het bezit van tanden in de( meeste )fossiele vertegenwoordigers .... Bovendien kloppen (meestal ) de lichaams-afmetingen ( en de verhoudingen ) ook al niet ...
Over het algemeen is een zoogdier een warmbloedig levendbarend dier dat zijn jongen voedt met moedermelk. Vanuit evolutionair perspectief mag je een vogelbekdier dus beschouwen als een zoogdier dat "nog niet helemaal af" is. Dit wil echter niet zeggen dat het dier niet over geavanceerde aanpassingen kan beschikken die hem in staat stellen te overleven in "modderige beken " bijvoorbeeld :.....Moderne vogelbekdieren ( waaronder ook fossiele vormen? ) zijn hoog -gespecialiseerde onderwater - jagers( op allerlei ongewervelden ?) die een soort elektro-locatie opspoor-systeem bezitten in hun snavel dienstig bij de voedselvergaring in troebel water ...
*Ook de moderne degenkrabben bezitten geen voldoende onveranderlijk bauplan in het fossiel archief , maar zijn wel al erg lang varianten op hetzelfde morfologisch conservatieve type sinds hun debuut in ????...? Animatie over de "vorm" van de degenkrabben --> http://www.blackwellpublishing.com/ridley/a-z/Stasis_c.asp ( Quick time noodzakelijk )
* Hetzelfde kan worden gezegd van de longvis - heeft de soort Neoceratodus forsteri (Australische longvis) een fossiel archief ? Natuurlijk hebben ze allemaal fossiele voorouders en familieleden, maar dat geldt voor alle gewervelden !
Ik vermoed dat een "LF" wordt beschouwd als een echt Levend Fossiel van zodra zijn stamlijn morfologisch strikt conservatief is gebleven gedurende het grootste deel van zijn bekende ( fossiel gedocumenteerde )evolutionaire geschiedenis in de geologische tijd (vanaf zijn opduiken in het fossielarchief tot op heden ) . Dat is echter een erg nutteloze onmogelijk te kontroleren ( en derhalve onbruikbare ) bepaling - want je kunt nooit weten of de daadwerkelijke stam-lijn altijd al sinds haar onstaan in het bezit was van een een onveranderlijk echt succesvol bauplan gelijk aan het huidige ( =dat dus niet evolutionair ontwikkeld werd en werd veranderd of gevarieerd ) Lintwormen hebben zelfs ook GEEN fossiele getuigenissen van hun evolutionaire geschiedenis en hun perfekt ( al altijd aanwezige ) ontworpen bauplan ; ze kunnen dus ook wel eens " kompleet , plotst en volmaakt geschapen " zijn , in de perfekte functionele vorm waaronder wij ze nu kennen ?
Rekening houdend met het voorafgaande zou men dus een 'levend fossiel' misschien kunnen definiëren als= ' een Genus - of een nog grotere taxonomische groep dan " soort " en/of ondersoorten - in het bezit van een morfologisch "archetype"(3) ( =oermodel) dat binnen een stamlijn van organismen , meer dan de normale gemiddelde geologische tijd 'behouden is gebleven . Maar zelfs dan, is het onmogelijk om zeggen wat "langer" is dan "gemiddeld" ....
Tyrannosaurs Rex , Kassuarisen en Thylacines zijn in dat geval dan slechts kortlevende uitzonderingen ? of andersom ? Hmmmm, wat is eigenlijk de "gemiddelde " norm ...?
(1) VEEL VAN WAT MENSEN ZIEN , IS WAT ZE MENEN/ VERWACHTEN TE ZIEN Bij nadere bechouwing is het duidelijk iets anders dan oorspronkelijk gedacht of "gezien" ... Maar alles wat wordt of werd ( ook vermeend ) "gezien" dient te worden verklaard ....
A) bij veel extante ongewervelde ( niet-verwante ) soorten is bijvoorbeeld ook een "gelijkvormige " lichaamsbouw het resultaat van"Mimycry" bijvoorbeeld imiteren zweefvliegen de lichaamsbouw (vespa) wespen ...Dat biedt de vliegen een serieus voordeel( in onze streken ) ...hun mogelijke predatoren ( vogels ) zien het verschil niet zo gemakkelijk en zullen waarschijnlijk veel minder het risico nemen een eventuele stekende wesp-achtige ( vespa) op te pikken ...
Verschil tussen wesp (bovenaan) en zweefvlieg (onderaan).
Mannetje, Garderen, 18-5-'05. Chrysotoxum cautum (Harris, 1776) grote fopwes
Zweefvliegen en wespen worden vaak met elkaar verward. Nochtans zijn ze, aan de hand van een aantal eenvoudige kenmerken, makkelijk van elkaar te onderscheiden. De naam zweefvliegen verwijst naar de manier waarop ze zich in de lucht verplaatsen. Ze vliegen met korte stukjes om dan plots te stoppen en ter plaatste te zweven. Een ander belangrijk kenmerk zijn de vleugels. Zweefvliegen hebben maar één paar vleugels terwijl bijen en wespen er twee paar hebben.
Overigens zijn en blijven voor veel leken "ongedierte " ,gewoon "ongedierte " Bijvoorbeeld : mieren zijn altijd mieren = dat er meer dan 8000 verschillende soorten zijn die onderling genetisch van elkaar geisoleerd blijven (NIET onderling kruisen )is blijkbaar voor velen niet interessant genoeg om weten ...en geenszins een onwetendheid die een sterke mening zou kunnen beletten .... zeker wanneer die meningsvorming nog eens vooroordelen bevestigd of gewoon gemakkelijk is ...
Trouwens de verwerving van de aangehaalde mieren -soorten -kennis wordt ook dikwijls beschouwd als nutteloze " beuzelarij " ; een soort bezigheidstherapie voor kindse mensen of onvolwassenen : Men weigert doodeenvoudig kennis op te doen over die vermelde feiten omdat men het een "tijdverspilling "noemt
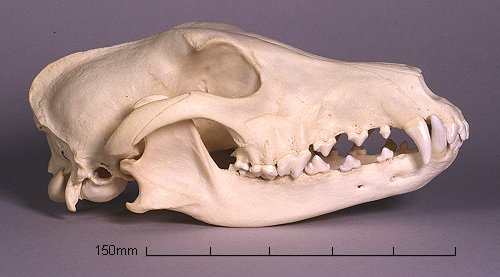
B) Ook convergente evolutie kan tot gelijkaardige morfologieen leiden zie bijvoorbeeld de schedels van buibelwolf / grey hound en grijze wolf
(Lateraal ) Dental Formula : 4.1.3.4 / 3.1.3.4 (Only 3 upper molars can be seen in this specimen, but the lower 4th molar is only partially erupted, and the animal must therefore have been immature.)
De significante verschillen ( bij het determineren van gevonden schedels bijvoorbeeld ) zijn voor de terzake specialisten- te ontdekken : die zijn georganiseerd in interdisciplinaire teams : in de systematiek, vergelijkende anatomie , de morfometrie (4) , de determinering , de paleontologie ( geassisteerd door de osteologen , geologen , genetici ..fylogenetici etc ) of te vinden in bijkomende analyses van eigenschappen van de vergeleken (fossielen en extante) soorten ; in bovenstaand voorbeeld : de formules voor het gebit ...
Het is nogal aanmatigend te denken dat onopgeleide niet-specialisten onzorgvuldige inschattingen van oppervlakkige en morfologische gelijkenissen (of verschillen ) " op het eerste zicht " , kunnen gebruiken om een conclusie te trekken aangaande "verwantschappên" en/of stamlijnen Dat geldt overigens niet alleen maar voor de inschatting van stasis in een SOORT op grond van "levende fossielen" en enkele" morfologisch gelijkaardige "(door creationisten geselekteerde )fossielen ....
(2) De twee extante brughagedissen-soorten ( Sphenodon punctatus (Gray, 1842) Sphenodon guntheri (Buller, 1877) Sphenodon diversum Colenso, 1885 (extinct) )zijn wel de enige in leven zijnde vertegenwoordigers van de reptielen-orde der rynchocephalia(= tegenwoordig bij voorkeur de orde Sphenodontia genoemd)ze bieden dus een uniek venster op een (bijna volledig ) uitgestorven groep ...
(3) *De term archetype ( in de paleontologie ) verwijst naar Richard Owen en T.Huxley *De creationisten noemen archetypes " het basisbouwplan van een baramin of een biblical kind " ze zien de gelijkenissen ( =homologieen ) tussen het ontwerp of de grond-plannen ( in feite plastische modellen waaraan wordt gerefereert door de morfometrie ) van verschillende soorten organismen als resultaten van én "micro-evolutie "( macro evolutie is gewoon geaccumuleerde " micro" - evolutie ) én " common design " (= ecomorphen & convergente evolutie ) * De term sloeg oorspronkelijk ( in de evolutionaire biologie ) op het morfologisch anatomisch gronddiagram van elk van de taxonomische Phyla
*omdat "archetype" ook veel "psychologische" en "magico-realistische" betekenissen heeft gekregen ( Jung ) is de term in hoge mate in onbruik geraakt...
* a) De huidigemorfologie /morfometrieis een zeer uitgebreide bundel wetenschappen (en zeer nuttige analytische instrumenten /hulpwetenschappen voor de vergelijkende anatomie / de systematiek, de cladistiek, de paleontologie etc ... ) geworden die zich niet meer alleen beperken tot het opstellen van de "grondplannen / grondmodellen van de phyla " ; http://www.digimorph.org/ b) de morfogenetische insteken ( in verband met de opeenvolgende verschijningsvormen en oorzaken van body-plans ) zijn een essentieel data -verzamelend onderdeel geworden van en voor allerlei morfogenetische studies die samen met de genetica ( = in het bijzonder sinds de ontdekking van de hox genen en de homeo-box) , deembryologie en de experimentele ontwikkelings-biologie het zeer belangrijke vakgebied van de evo-devo (= evolutionary development biology )hielp onstaan ....
(Artikel van Gerdien De Jong ) HET DIER ; EEN BOUWPAKKET
....In de aanvaarding van het idee van evolutie als afstamming onder verandering hebben de anatomie en ontwikkelingspatronen ( body- plans/ Morfogenesis )van dieren een grote rol gespeeld.
Maar daarna gingen de evolutie- en ontwikkelingsbiologie elk hun eigen weg.
Een nieuw raakvlak ontbrak zolang de genetica binnen de embryologie geen rol speelde. Embryologen zagen een moeilijkheid in de genetische bepaling van ontwikkeling: hoe zou lineair DNA kunnen coderen voor driedimensionale structuren?
In 1980-1984 werd de genetica van de embryonale ontwikkeling ontraadseld; in 1984 bleek dat de embryonale ontwikkeling van zeer verschillende dieren gebruikmaakte van dezelfde genen. Als gevolg van deze genetische revolutie in de ontwikkelingsbiologie ontstond het vakgebied Evolutionary Developmental Biology, evo-devo.
EVOLUTIE BETEKENT ALTIJD DAT DE ONTWIKKELING VAN DIEREN VAN EI TOT VOLWASSENE VERANDERT, EVOLUEERT. ( lees verder op ---> Het dier een bouwpakket (Tsjok45 )
De 'genetische gereedschapskist'
Sinds Walter Gehring weten we dat bepaalde hox genen ( bijvoorbeeld het "no-eyes gen" ) onderling verwisselbaar zijn bij verschillende diergroepen :
....dezelfde genen worden steeds weer gebruikt, zodat belangrijke evolutionaire veranderingen niet noodzakelijk belangrijke genetische veranderingen nodig hebben. Er is complexiteit in het tempo en de modus van evolutie. Er zitten veel verschillende patronen in micro-evolutionaire gebeurtenissen. Micro-evolutie en macro-evolutie zijn voortdurende processen...
4) Twee belangrijke onderdelen van de morfometrie :
- Allometry ( T. Hyxley ) bijvoorbeeld ; EQ ( encefalisatiesuotient = de verhouding tussen hersenmassa,en lichaamsmassa ....
- D'Arcy Wentworth Thompson transformations De( soorteigen) morfologische varianten op een "grondplan geprojecteerd op een raster/coordinaten " ( wordt ook gebruikt in 3D computer tekeningen )
Johan Gielis is de bedenker van de superformule, die hij ontdekte bij een onderzoek naar modellen die plantenvormen beschrijven. Omdat niet bekend was of en zoja, welke wiskundige regels gelden voor de vorm van natuurlijke organismen, construeerde Gielis een wiskundige formule. Hierbij nam hij als uitgangspunt de "superellips" van de 19e eeuwse Franse wiskundige Gabriel Lamé. Dit resulteerde uiteindelijk in de superformule die het mogelijk maakt verschillende abstracte en natuurlijke vormen zoals planten, schelpen, cirkels, ruimte-tijdmodellen en kristallen, met één enkele formule te beschrijven.
"From the contemplation of plants, men might first be invited to mathematical inquiries (N. Drew, 1682)"
In the past, the idea of optimization for some sophisticated purpose seemed the only conceivable explanation for the level of complexity in biology.
Take the example of fish which manifest so many beautiful colors. One Darwinian explanation would be that these colors improved survival through either allowing the creatures to evade being hunted by blending with the environment or shocking the predator with their brilliant, contrasting colors.
In other words, every one of these colors and patterns evolved for a purpose. But then why do all the multi-colored fish have the same exact internal structures while having such radically different patterns on the outside? Wolframs beguilingly simple explanation is that the most visually striking color differences have almost nothing to do with natural selection, but are reflections of completely random changes in underlying genetic programs.
On the other hand, the vital features such as the internal organs have changed only quite slowly and gradually in the course of evolution because those are precisely the ones molded by natural selection.
The range of pigmentation patterns on mollusc shells (picture) correspond remarkably closely with the range of patterns that are produced by simple randomly chosen programs based on cellular automata. There are already indications that such programs are quite short. One of the consequences of a program being short is that there is little room for inessential elements and any mutation or change in the program, however small, will tend to have a significant effect on at least the details of patterns it produces. Or biologic systems should be capable of generating essentially arbitrary complexity by using short programs formed by just a few mutations (ANKS).
But if complexity is this easy to get, why is it not even more widespread in biology? The answer is natural selection, which can achieve little when confronted with complex behavior. There are several reasons for this. First, with more complex behavior, there are huge numbers of possible variations. Second, complex behavior inevitably involves many elaborate details, and since different ones of these details may happen to be the deciding factors in the fates of individual organisms, it becomes very difficult for natural selection to act in a consistent and definitive way. Third, whenever the overall behavior of a system is more complex than its underlying program, almost any mutation in the program will lead to a whole collection of detailed changes in the behavior, so that natural selection has no opportunity to pick out changes which are beneficial from those which are not (ANKS).
In the picture here, one can see the shell shapes generated by the simple model and those actually found in nature. The computerized array shows systematic variations of just two parameters.
Progress in science occurs when a new order is discovered which can unite seemingly unrelated facts.
ANKS has achieved just that by showing that it is possible to get complexity from simple programs in any system.
The job of a scientist in the conventional sense is to understand how things work and not why.
Wolfram has combined the two by asking not only how evolution occurs, but also the meaning of it all. And the answer is terribly disturbing. No meaning at all.
So why do higher organisms exist at all? My guess is that it has almost nothing to do with optimality, and that instead it is essentially just a consequence of strings of random mutations that happened to add more and more features without introducing fatal flaws (ANKS p398).
De handleiding is ingewikkelder dan die van een bouwmarkt of Ikea, maar essentieel zijn ook hier niet de onderdelen maar de manier waarop ze aan elkaar geschakeld worden. Evolutie is de ontwikkeling van toepassingen.
Hoe verschillend zijn dieren van elkaar?
Hebben alle dieren, van kwallen tot mensen, in principe hetzelfde bouwplan? Of bestaan er fundamentele verschillen tussen de bouwplannen van elke diergroep? Het antwoord op deze vraag is van groot belang voor de evolutiebiologie. Als er sprake is van een gemeenschappelijk grondplan, dan betekent dit dat er in de evolutie voornamelijk gevarieerd wordt op eenzelfde thema. Elk dier bezit dan zijn eigen interpretatie van dat grondplan, met gebruik van dezelfde genen. Maar als er fundamentele verschillen bestaan tussen de bouwplannen, dan moeten telkens, voor elke groep dieren, volledig nieuwe genen uitgevonden worden. .....
In 1980 werd duidelijk dat de fruitvlieg acht genen voor homoeotische mutanten heeft. Deze genen bepalen in het embryo de functie (identiteit) van de al gevormde segmenten, werkend van voor naar achter in dezelfde volgorde als ze in het DNA liggen. De grote doorbraken in de daaropvolgende jaren kwamen voort uit moleculair onderzoek aan deze acht genen. Ze bleken een stukje DNA gemeenschappelijk te hebben (1984).
Dit DNA leidde tot een eiwit met als belangrijkste functie de regeling van andere genen. De acht genen voor homoeotische mutanten nu Hox-genen geheten leveren het gereedschap voor het aan- of uitzetten van schakelaars.
De stand van deze schakelaars bepaalt of een bijbehorend gen aan- of uitstaat. De uiteindelijke functie/vorm van een segment langs de voor-achter as wordt bepaald door de specifieke aan/uit combinatie van de Hox-genen,( = de master-switches ) op basis waarvan andere genen aan- of uitgezet worden.
In 1984 vond men dat de regenworm, de pad, de kip, de muis en de mens dezelfde Hox-genen hebben, die in dezelfde volgorde werken om de identiteit van de onderdelen langs de voor-achter as aan te geven (zie Figuur 1).
Figuur 1. Expressie van de genen pax 6 (maak hier oog), otx en emx (maak hier hersenen) en de Hox-genen langs de voor-achter as, bij embryos van de fruitvlieg, de muis en de eikelworm, een primitief beest uit de Deuterostomia, de groep waartoe de gewervelde dieren behoren.
Naast Hox werden vele andere genen voor gereedschapseiwitten gevonden. Deze geven commandos als maak hier oog, maak hier hart, maak hier brein.
Wat voor een oog, hart of brein het wordt, hangt af van het dier. Hersenen bij zoogdieren en insecten hebben weliswaar een totaal verschillende anatomie, maar de achterliggende genen zijn hetzelfde.
Hoe maak je nu een dier?
Men neme een beginnend embryo. Specificeer de voor-achter as en de onder-boven as. Zet coördinaten langs beide assen. Specificeer aan de hand van dit coördinatenstelsel waar een derde as aangrijpt: de van-binnen-naar-buiten as. Zet een eigen coördinatenstelsel op in de uitstulping. Specificeer nu de onderverdeling van het lichaam, de identiteit van de onderdelen. Dit alles is wat ontwikkelingsgenen doen: de plaats in het embryo bepalen en die plaats een identiteit geven, onder andere met behulp van Hox-combinaties. De zeven segmenten van de spinnenkop werken bijvoorbeeld met Hox 1-6; de negen segmenten van het spinnenachterlijf met Hox 7-10. De zes segmenten van de insectenkop werken met Hox 1-2, 4-5; de drie segmenten van het borststuk met Hox 7 en de acht segmenten van het achterlijf met Hox 8-10. Insecten en spinnen vertonen verschil in bouw bij dezelfde elementen van het bouwplan.
Hoe maak je nu iets nieuws? Door het schakelschema te veranderen of door onderdelen te hergebruiken. Zo komen de vleugels van insecten genetisch overeen met de kieuwen van kreeften.
Schakelaars hergebruiken is een andere mogelijkheid. Het gen Distal-less wordt aangeschakeld in het embryo van insecten voor de plaatsbepaling van poten en monddelen: in de poot voor de klauwtjes, maar ook voor de aanleg van de vleugel en voor de plaats van de toekomstige vleugelrand. Vlinders hebben daarbij nog een mogelijkheid voor Distal-less: dit aangeschakelde gen vertelt waar in de vleugel een oogvlek moet komen (Figuur 2).
De ontwikkeling van een nieuwe tentakel aan de monddelen van Yuccamotten gebruikt ook Distal-less voor de plaatsbepaling.
Soms lijkt er niet zoveel nodig voor een nieuw bouwplan. Vleermuizen evolueerden omstreeks 53-50 miljoen jaar geleden, op het hoogtepunt van de diversiteit in insecten. Vleermuizen zijn een succesvolle groep: één op de vijf zoogdiersoorten is een vleermuis. De groep dankt dit succes onder andere aan de goede vleugels. Die zijn bij alle vleermuizen hetzelfde gebouwd, namelijk door een verlenging van de vingers. Tussen vingers, lijf en achterpoten is een vlieghuid gespannen. Zon vlieghuid is op zich niet bijzonder, een aantal niet-verwante zoogdieren heeft er ook een, zoals de sugarglider, de vliegende eekhoorn en de vliegende lemur. Het grote verschil tussen de vliegende lemur en de vleermuis zit in de lange vleermuisvingers.
Die ontwikkelen zich als volgt. De hand van de vleermuis wordt op vergelijkbare wijze aangelegd als de hand van de muis. Wanneer het kraakbeen zich begint te vormen, zijn de vingers van muis en vleermuis nog identiek. Groei en differentiatie van de vingerbotjes staan onder controle van een aantal genen, onder andere Bmp2. Dit gen is actiever in de vleermuis, wat leidt tot een snellere groei en langzamere differentiatie van het weefsel, dus tot langere vingers. Het schakelsysteem tussen muis en vleermuis is veranderd, zodat het gen Bmp2 bij vleermuizen sterker werkt. Dit is het cruciale verschil, een vernieuwing die heeft geleid tot de snelle verspreiding van vleermuizen. Geen grote verandering in de ontwikkeling, wel een grote evolutionaire vernieuwing.
Figuur 2. Distal-less schakelaars: een extra schakelaar voor oogvlekken op de vleugel van een vlinder.
Is ID de wetenschappelijke (sic) uitleg voor creationiisten 150jaar Darwinisme... De evolutietheorie gewikt en gewogen David sorensen en revolutietheorie lalala DE VERRIJZENIS VAN DE JONGE VERDRONKEN KOE » Reageer (21) ID TOVERDOOS OVER SLECHTE ID-EETJES ,RESISTENTIE & TOVERDOCTORS c) OEC d) YEC
Een oeroud spoor Denisova : ZUID SIBERIË DENISOVA - mens
DMANISI AAP OF MENS DE OUDSTE ? » Reageer (1)Nakalipithecus nakayamai (<) Alweer eentje ? ..... De tand des tijds / Heidelberg-mens DE HEUPEN VAN EVA CASABLANCA MAN /erectus Hobbit is aparte soort ? » Reageer (2) NOG EEN BENDE BIJTERS POLONAISE met neanderthaler TANDEN UIT DE QESEM GROT Neanderthaler genoom Mitochondriale Genenkaart van Neanderthaler Xuchang mens
b)Biologie 1.-(EVOLUTIE ) NAS / IM Document 2008 (2) NAS / IM Document 2008 ( 1)
Cambrium & precambrium AVALON GABONESE chips
Evolutie in actie Opmerkelijk snelle adaptaties bij kroatische ruine-hagedissen:
EEN PLUIM VOOR CHINA Geef eens een pootje Pluimgewicht KLEIN DUIMPJE & DE REUS &VEREN Opnieuw gevederde Maniraptor Similicaudipteryx. Aerosteon riocoloradensis
Krokodillen
PAKASUCHUS KAPILIMAI Prestosuchus chiniquensis Terug naar zee viseter
Eritherium azzouzorum LJOEBA Darwinius masillae VLIEGENDE KATTEN ? Vleermuizen ALWEER EEN BELANGRIJK STUK UIT DE LEGPUZZEL Zee-zoogdieren ; Van de wal in de visgronden Indohyus / walvisevolutie Zeehonden-evolutie
*Blauwe en groene ( =nederlandse)teksten zijn meestal aanklikbare links *Engelse Wikipedia teksten verwijzen in de linkerkolom naar verschillende niet-engelse versies van het wikiartikel *Blauwe teksten tusssen "" , zijn voornamelijk ( gedeeltelijke)citaten afkomstig van mensen met andere meningen
Evolutie / Charles Darwin aan de basis: SELECTIE NATUURLIJK DE LEVENSBOOM UITSTERVEN DEEP TIME Biogeografie SEKSUELE SELECTIE Al jaren lang samen CO-EVOLUTIE De economie van de natuur Geleidelijke veranderingen
Unintelligent design:
IP Theorie Ons gebit , De appendix, Geboorte, De weg van het zaad , Nervus Laryngeus Recurrens, DNA , 75 mistakes , Het boek
Unintelligent design (1): Rechtop lopen Tomaso over het oog , oogspieren , Unintelligent Design (5): ZWEETVOETEN Unintelligent design (6): Vitamine C Unintelligent design (8): Kuitspieren
 Unintelligent design (1): Rechtop lopen
Unintelligent design (1): Rechtop lopen








 Verschil tussen wesp (bovenaan) en zweefvlieg (onderaan).
Verschil tussen wesp (bovenaan) en zweefvlieg (onderaan).
)





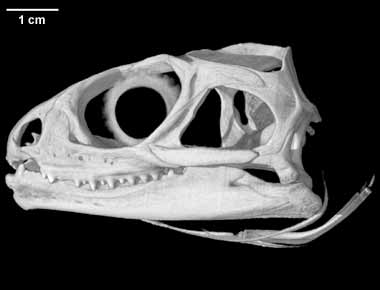 <http://www.digimorph.org/specimens/Sphenodon_punctatus/adult/
<http://www.digimorph.org/specimens/Sphenodon_punctatus/adult/
















 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}
{kind=link}