Dit blog is een onderdeel van EVODISKU WAT IS DE BEDOELING EN WAAR STAAT DIT BLOG VOOR
****Wie meent dat alles inmiddels wel over de evolutietheorie gezegd is en dat de discussie gesloten kan worden, ziet over het hoofd dat de wetenschap niet stil staat.
***Wie meent dat inmiddels het creationisme definitief het pleit heeft verloren en dat de discussie gesloten kan worden , ziet over het hoofd dat het "creationisme" is geevolueerd ( en zal evolueren ) in nieuwere mimicrytische vormen( meme-complexen ) zoals bijvoorbeeld het ID(C)
***Dit blog is speciaal opgezet om de aktualiteit binnen het evolutie-creationisme debat te volgen en van kommentaren te voorzien ... waartoe de lezers zijn uitgenodigd bij te dragen ...
Let echter wél op het volgende :
"Je bent een rund als je hier met religie stunt " ....
19-03-2008
KWASTEN
Levende fossielen (1) :
"Levende fossielen" (= volgens de creationisten , de pop wetenschap en de meeste kranten van jaren geleden ) zijn niet hetzelfde als het begrip " levende fossielen "(= volgens de Paleontologen , systematici en de zoologen )
Er zijn geen miljoenen jaren oude fossiele exemplaren bekend , die ( zelfs maar morfologisch )volkomen identiek zijn aan nu nog levende (=extante) soortenen die men vrij generaliserend ( en in het ondertussen ingeburgerde taalgebruik ) "levende fossielen "noemt ... Dit werd al een tijdje geleden goed onderbouwd door een voormalig YEC-creationist die ondertussen "christelijk" wetenschapper( theistische evolutionist --> http://www.dailykos.com/story/2006/1/21/111430/597 ) is geworden ....
LIVING FOSSILS --THERE ARE NONE
Copyright 2003 G.R. Morton
This can be freely distributed so long as no changes are made and no charges are made.
( hier volgt een voorlopige vertaling van een passus uit dat werk )
" ...Toen ik (nog) een jong-aardecreationist was, geloofde ik in levende fossielen. Dan begon ik de details te bekijken en kon ik geen één enkel levend ` fossiel ' vinden dat feitelijk identiek was aan de oude vorm ( gedocumenteerd in de daadwerkelijke fossielen ) . Deze levende fossielen zijn : ( volgens de creationisten =) "vermoedelijk oude en lang uitgestorven schepselen die plotseling en onverwacht levend te voorschijn komen ( ontdekt werden) in de moderne wereld."(Whitcomb and Morris, THE GENESIS FLOOD, p. 176) Maar het blijkt, dat er GEEN "levende fossielen "te vinden zijn , tenminste als men daarmee bedoelt : een levende plantensoort of levende diersoort die morfologisch gelijk is aan (in voldoende mate gevonden exemplaren van ) fossiele tegenhangers . Dergelijke " levende fossielen" zijn vrij verschillend van de vooropgeschoven 'oude vormen.' "
---> Meer weerleggingen (1) (Tsjok45)zie ook (1)
Het levende fossiel bij uitstek is uiteraard Latimeria = de Coelacanth:
Een roofvis van 1,30 - a meter ,behorend tot een oeroude familie spiervinnige vissen, de Actinistia. Latimeria is hiervan het enige nog levende geslacht( genus) . In tegenstelling tot wat maar al te gauw beweerd of sterk gesuggereerd /geconcludeerd wordt , zijn huidige vertegenwoordigers van Latimerianiet bekend van een 380 miljoen jaar oud fossiel. Het meest op Latimeria lijkende fossiel is Macropoma uit het late Krijt. *
De laatste bekende spiervinnige fossielen (van de Coelacanthiformes ) stammen ,tot op heden , uit het laat of boven-Krijt van Zuid- Frankrijk ( > 2005 (Tsjok45) , maar tot 1938 werd gedacht dat de Actinistia sindsdien verdwenen waren
Dat het zo lang duurde tot wetenschappers deze vis" levend " hadden gevonden is nog niet zo héél vreemd, de hedendaagse Coelacanth leeft in diep water, en houdt zich lange tijd schuil in onderzeese grotten. Het duurde pas tot 1952 tot er een tweede exemplaar werd gevonden. Hierna daalden biologen in een duikboot af op tweehonderdtwintig meter en zagen toen pas voor het eerst levende Coelacanthen. Ze maakten met hun vier vlezige vinnen een soort loopbewegingen in het water en marcheerden zo door de duisternis.
Volgens Ichtyologen waren deze Coelacanthen eigenlijk helemaal 'verdwaald'. Als voorouders van de tetrapoden zouden ze eigenlijk eerder aan de oppervlakte moeten leven, in zoet water. Fossiele spiervinnigen zijn inderdaad gevonden in zoetwater afzettingen, maar deze Coelacanthen leven in de diepzee, ver van het droge. Echter , tegenwoordig worden de coelacanthidae niet meer beschouwd als devoorouderlijke groep van de tetrapoda-lijn ...Dat is nu de groep der Rhipidistia geworden
Mogenlijk hebben de spiervinnigen vanaf het Krijt hun tehuis aan de oppervlakte opgegeven voor een leefgebied in de grotten van de diepzee. Zulke grotten zijn slechte plaatsen voor fossielisatie, dus logisch dat paleontologen vanaf dat moment hun spoor kwijtraakte. Een andere factor is dat de Coelacanthen sindsdien erg zeldzaam zijn geworden. Tegenwoordig zijn er ( voorlopig slechts ) twee species bekend; L.chalumnae, gevonden nabij Zuid-Afrika, en L.menadoensis uit Indonesië.
Matt Friedman, (een afgestudeerde student aan de Universiteit van Chicago in de VS, )ontdekte in 2005 een uniek fossiel dat een beter begrip mogelijkmaakt over de evolutie van de vinnen van de huidige coelacanthen De ontdekking werpt vooral licht op een vraag die al lang veel wetenschappers bezig hield : " ....hoe is het mogelijk dat de symetrische vinnen van deze vis, ( waarvan gedacht werd ) zo nauw verwant (te zijn )aan landdieren, ontwikkeld zijn tot de assymetrische handen en voeten van tetrapoden ? "
De in 1938 ontdekte extante coelacanth werd in de media en de popwetenschap erg vlug geprezen als een " echt " levend fossiel. Voordien dacht men dat de coelacanthen meer dan 65 miljoen jaar geleden kompleet waren uitgestorven . De twee paar ventrale vinnen waren bijzonder belangwekkend want ze werden verondersteld te zijn geëvolueerd vanuit dezelfde structuren waaruit ook de ledematen van landdieren zich ontwikkelden .
De opmerkelijke vinnen van latimeria vormde een raadselachtige puzzle : in tegenstelling tot de handen en voeten van landdieren waren ze symmetrisch.
Recente fossiele vondsten hadden echter voor 2005 al aangetoond dat handen en voeten zijn geëvolueerd uit uitgestorven voorouderlijke vissen met asymmetrische vinnen: de voorouders der tetrapoden werden van toen af geplaatst in de groep der Rhipidistia
Maar de vraag hoe de huidige coelacanth haar symmetrische vinnen dan wel verkreeg ,was gebleven.
Verloren de coelacanthus - vinnen hun oorspronkelijke asymmetrie? m.a.w. waren zij om te beginnen ( in een basis oer-vorm) al asymmetrisch? Hoewel veel verschillende soorten coelacanth fossiel zijn gevonden, bewaarde geen van hen de cruciale skelet -details , waarschijnlijk omdat deze grotendeels gemaakt zijn van kraakbeen - en dat is bijna nooit bewaard gebleven ...
Dat alles veranderde in 2005, een belangrijk jaar voor het coelacanthen onderzoek
Friedman was op fossielenjacht in het puin aan de voet van een klif in Wyoming, toen hij over een interessant stuk rots, struikelde . Wat hij aanvankelijk voor een plant fossiel hield bleek het enige ( tot nu toe bekende )fossiel van een voorouder van de coelacanthen- vin .
"Het was echt een ongelooflijk bof ", zegt Friedman. Het 10 - centimeter - fossiele viel van ten minste 70 meter hoog van de klif omlaag , zonder te verbrijzelen - de vondst bevat de schouder en vin- botten . ze zijn goed gefossiliseerd omdat ze minder kraakbeen bevatten dan de vinnen van de levende coelacanth. De botten zijn verbonden op een manier die kenmerkend is voor coelacanthen en lieten Friedman en zijn collega's toe in het fossiel een voorouder te herkennen :
Shoshonia arctopteryx , had asymmetrische vinnen. Dit geeft aan dat de levende coelacanthen de symmetrie later hebben ontwikkeld
Dat Shoshonia en de huidig nog levende coelacanths verschillen is niet verwonderlijk ( en ondanks de creationistische onzin ) - coelacanthen hebben immers een evolutionaire geschiedenis van minstens 400 miljoen jaar achter de rug .
Maar het is ook een lesje voor degenen die de ontwikking van bepaalde onderdelen van organismen bestuderen .
"De asymmetrie in onze eigen gepaarde ledematen is in feite een primitief kenmerk" zegt Michael Coates van de Universiteit van Chicago, VS.
Terwijl de coelacanth deze eigenschap niet heeft gehandhaafd is dat wel het geval bij andere, meer primitieve levende vissen . ondanks het feit dat de coelacanth een zeer verre maar nauwe verwant is van landdieren, "Kan men nu beter onderzoek doen bij lepelsteuren , steuren , en haaien"
Sharks (like this Great White off the coast of Guadalupe Island, Mexico) have the recipe for fingers in their genetic cookbook, researchers have found. (Credit: iStockphoto/Keith Flood)
KWASTVINNIGEN 8Families /7 uitgestorven
Coelacanthiformes xml:namespace prefix = o ns = "urn:schemas-microsoft-com:office:office" />
Kwastvinnigenbestaan al (minstens ) vanafhet Devoon (410 MY): De grote vormenrijkdom dateertvan het Trias: de laatste fossielerestanten( van de familie latimeridae) zijn afkomstiguit het krijt.(-90 MY) .. Daarom worden de hedendaagse coelacanthen'levende fossielen' genoemd . dehuidige coelacanthenzijn echter leden van het genus Latimeria en niet van hetlaatst nog bekende fossielegenus / uit het krijtMacropoma ( beide genera behorenwél tot dezelfdefamilie Latimeridae)
Kwastvinnigenzijn beenvissen en vormen samen met de longvissen totde Sacopterygii. ( spiervinnigen ) Kwastvinnigen zijn vissen met borstvinnen en buikvinnen, die worden gesteund door een aantal scharnierende skeletelementen. De leden van degroepder spiervinnigen hebben als belangrijkegemeenschappelijke kenmerk ;vier gepaarde , vlezigeen gespierdevinnen.
Het zijn de enige extante recente gewervelde dieren met een scharnierpunt in de schedel. ( ook fossiel aantoonbaar ) Door dit scharnier kan de bovenkaak omhooggetrokken worden. Waarnemingen bij recente Latimeria , hebbenaangetoonddat deze kwastvinnigenvolgens de bewegingsprincipes der gewerveldelanddieren (Tetrapoden) zwemmen(= alternerend synchroon )
Ze leven van kleine vissen. De hedendaagse Coelacanthen kennen een inwendige bevruchting. De eieren komen uit in het lichaam van de moeder, waar de jongen zich nog enige tijd ontwikkelen voordat ze geboren worden Dooierzakjes van uitgekomen eitjesen (waarschijnlijk het teveel aaneitjesen larven ) worden verteerd door de moeder
De kwastvinnigen worden onderverdeeld inRhipidistia(.bijvoorbeeld: Osteolepis)die worden beschouwd alsde voorlopers groep waaruit de tetrapoden zich ontwikkelden .;enCoelacanthidea (bijvoorbeeld de zogenaamde. CoelacanthusAgassiz: heteerste holotype Coelacanthus granulatus( = granulosus )
Onveranderlijk sinds miljoenen jaren? Men bedoelt met levende fossielen organisen met een ietwat prehistorisch uiterlijk, of waarvan het fossielenbestand ons kan vertellen dat deze al rondliepen -of zwommen in de tijd van de dinosauriërs of een ander ver verleden. Vaak hoor je dan de populaire kreet; "onveranderlijk gebleven sinds vele miljoenen jaren". Maar is dit ook zo? Het antwoord is dat je enkel met behulp van een hedendaags exemplaar, en de fossiele resten van een prehistorisch exemplaar niet kunt vaststellen dat er geen evolutie heeft plaatsgevonden. Niet alle evoluties spelen zich namelijk af aan de buitenkant van een organisme, er kunnen ook veranderingen optreden in het gedrag of instinct, het immuunsysteem of de (schud)kleur en interne organen (die fossiliseren immers niet.). Kortom, een fossiel dat sprekend lijkt op een modern exemplaar is geen bewijs van onveranderlijkheid, want deze stelling baseert zich slechts op het uiterlijk dat je van het fossiel kan afleiden. De stelling "onveranderlijk" komt hiermee al op losse schroeven te staan. Toch worden dergelijke dieren door bepaalde personen gebruikt als bindmiddel voor hun hypothese dat alle bekende huidige en prehistorische dieren rond de zelfde tijd zouden moeten hebben geleefd. Ze hopen hiermee aan te tonen dat dieren die als 'prehistorisch' worden omschreven soms nog gewoon vandaag de dag leven en dat er geen evolutie heeft plaatsgevonden.
Levende fossielen Een introductie tot enkele zogenoemde levende fossielen:
Een artikel van Tim Spaan.
Onveranderlijk sinds miljoenen jaren? Letterlijk genomen is een levend fossiel zoiets als een vierkante cirkel, het spreekt zichzelf tegen. Fossielen zijn immers de versteende resten van al lang afgestorven organismen. Maar men bedoelt er uiteraard dieren mee met een ietwat prehistorisch uiterlijk, of waarvan het fossielenbestand ons kan vertellen dat deze al rondliepen -of zwommen in de tijd van de dinosaurirs of een ander ver verleden. Vaak hoor je dan de populaire kreet; "onveranderlijk gebleven sinds vele miljoenen jaren". Maar is dit ook zo? Het antwoord is dat je enkel van een hedendaags exemplaar, en de fossiele resten van een prehistorisch exemplaar niet kunt vaststellen dat er geen evolutie heeft plaatsgevonden. Niet alle evoluties spelen zich namelijk af aan de buitenkant van een organisme, er kunnen ook veranderingen optreden in het gedrag of instinct, het immuunsysteem of de (schud)kleur en interne organen (die fossiliseren immers niet.). Kortom, een fossiel dat sprekend lijkt op een modern exemplaar is geen bewijs van onveranderlijkheid, want deze stelling baseert zich slechts op het uiterlijk dat je van het fossiel kan afleiden. De stelling "onveranderlijk" komt hiermee al op losse schroeven te staan. Toch worden dergelijke dieren door bepaalde personen gebruikt als bindmiddel voor hun hypothese dat alle bekende huidige en prehistorische dieren rond de zelfde tijd zouden moeten hebben geleefd. Ze hopen hiermee aan te tonen dat dieren die als 'prehistorisch' worden omschreven soms nog gewoon vandaag de dag leven en dat er geen evolutie heeft plaatsgevonden. Dan stel ik mij opnieuw de vraag: is dit ook zo? Om te beginnen neem ik eerst eens wat zogenoemde levende fossielen onder de loep:
Latimeria de Coelacanth Het levende fossiel bij uitstek is uiteraard Latimeria de Coelacanth (Spreek uit: Sie-la-kant en niet Koelakant.), een roofvis van iets minder dan twee meter die behoord tot een oude familie, de Actinistia. Latimeria is hiervan het enige nog levende geslacht. In tegenstelling tot wat maar al te gauw beweerd wordt is Latimeria niet bekend van een 380 miljoen jaar oud fossiel. Het meest op Latimeria lijkende fossiel is Macropoma uit het laat Krijt.
De bewering dat Coelacanthen miljoenen jaren lang onveranderlijk zijn gebleven is dus sowieso onjuist. Klik hier om de verschillen te zien tussen Latimeria en Macropoma. Let ook op het verschil in grootte (plaatjes zijn niet op schaal). De tegenwoordige Coelacanth is drie maal groter dan de jongste soort in het fossielenbestand. Tot 1938 werd gedacht dat de familie Actinistia verdwenen was sinds het Krijt, en dat Macropoma de laatste Coelacanth was die de wereld had aanschouwd. Dat het zo lang duurde tot wetenschappers de moderne Coelacanth hadden gevonden is nog niet zo hl vreemd, de hedendaagse Coelacanth leeft in diep water, en houdt zich lange tijd schuil in onderzeese grotten. Het duurde pas tot 1952 tot er een tweede exemplaar werd gevonden. Hierna daalden biologen in een duikboot af op tweehonderdtwintig meter en zagen toen pas voor het eerst levende Coelacanthen. Ze maakten met hun vier vlezige vinnen een soort loopbewegingen in het water en marcheerden zo door de duisternis. Men zou denken dat Coelacanthen, die ten slotte betrokken zijn geweest bij het ontstaan viervoeters, eerder dicht aan de oppervlakte zouden hebben geleefd in plaats van in de diepzee. Maar dat is dus niet het geval. Blijkbaar hebben de vissen vanaf het Krijt hun tehuis aan de oppervlakte opgegeven voor een leefgebied in de grotten van de diepzee. Zulke grotten zijn slechte plaatsen voor fossilisatie, dus logisch dat paleontologen vanaf dat moment hun spoor kwijtraakte. Een andere factor is dat de Coelacanthen sindsdien erg zeldzaam zijn geworden. Tegenwoordig zijn er twee species bekend; L.chalumnae, gevonden nabij Zuid-Afrika, en L.menadoensis uit Indonesi. Onderzoeken naar de Coelacanth brachten overigens aan het licht dat deze niet de nauwste verwanten van viervoeters zijn. Die plaats is voorbehouden aan de longvissen, zo wezen testen met het mitrochondriale DNA van longvissen uit. Desondanks vertoond het DNA van Latimeria nog wat kenmerken die aan verwantschap met viervoeters doen herinneren, zo is het bloedhemaglobine van de Coelacanth gelijkaardig aan dat van kikkervisjes, maar niet aan dat van volwassen kikkers. Bovendien werd ontdekt dat het de enige nog levende gewervelde is met een intercraniale verbinding, wat ooit gemeenschappelijk was voor kikkers. Door deze verbinding kan de bovenste kaak omhoog worden gebracht en tegelijkertijd de onderkaak naar beneden, wat het volume van de bek enorm vergroot. Evenals haaien ontwikkelde de Coelacanth een spiraalvormig spijsverteringsstelsel die de oppervlakte van de darm, waarmee de passerende voedselstoffen in aanraking komen, aanmerkelijk vergroot. En net als haaien heeft het dier ook een kraakbeenskelet, met in de wervelkolom een begin van verbening. De kop, tanden en schubben zijn inmiddels al geheel van been.
Degenkrabben (bv. Limulus polyphemus)
De voorouders van degenkrabben ontstonden 360 miljoen jaar geleden, maar deze hadden alleen nog een vage degenkrabachtige vorm. Echte degenkrabben ontstonden pas in het late Jura.Degenkrabben vervulde tijdens het Jura en Krijt de rol van krab, totdat ze na het Krijt werden vervangen door de moderne krabben. Hoewel ze het daarna nog 65 miljoen jaar hebben uitgehouden is de degenkrab vergeleken met vroeger nu zeer zeldzaam. Vandaag de dag zijn er 3 geslachten die voorkomen aan de westkust van Noord-Amerika, India en oost-Azi.
Sphenodon punctatus, de Tuatara.
Hij ziet er uit als een hagedis, maar is het eigenlijk niet. Hagedissen behoren tot de orde der Squamata, maar de Tuatara behoort tot een veel oudere familie die de Sphenodontia wordt genoemd. De Tuatara leeft in Nieuw-Zeeland, en is het enige nog levende lid van de Sphenodonten, maar vroeger heeft deze familie vele verschillende soorten gehad. Dat deze zijn uitgestorven komt doordat ze de concurentiestrijd met de ware hagedissen niet aankonden. Deze laatsten beschikken over betere aanpassingen zoals een flexibele kaak, en hebben in de hele wereld de oude Sphenodonten sinds ongeveer 100 miljoen jaar vervangen. Behalve in Nieuw-Zeeland dus, waar de Tuatara, die voornamelijk leeft van Weta's (grote sprinkhanen) blijkbaar niet direct concureerd me de Nieuw-Zeelandse hagedissen (er leven daar alleen Gekko's en Skinken). Maar waarom is dit dier nu een levend fossiel? Laat ik voorop stellen dat van Sphenodon punctatus er gn fossielen zijn gevonden, geen oude en geen recente. Bijgevolg kan op geen enkele manier worden bewezen (kan evengoed al niet, zie inleiding) dat dit dier sinds miljoenen jaren onveranderlijk is gebleven. Hoewel het niet zo waarschijnlijk is kan de Tuatara van allerlij evoluties hebben doorgemaakt waar we niks van weten. De enige reden waarom men het een levend fossiel noemt is vanwege die oude familieband, maar daarom is het nog wel een gewoon modern dier.
Waarom zulke oude familieleden?
Dit zijn enkele van de voornaamste, maar niet alle, voorbeelden. Nader beschouwd zien we dat ook waar het morfologische verandering betreft, de onveranderlijkheid niet opgaat, de huidige Coelacanth verschilt aanzienlijk van de prehistorische. Ook van de Tuatara kunnen we niet zeggen dat deze miljoenen jaren lang onveranderlijk is gebleven en ook de degenkrabben hebben een ontwikkelingsgeschiedenis achter zich. En dit geldt voor vrijwel alle levende fossielen, het zijn moderne dieren, wiens familie miljoenen jaren geleden is ontstaan.
In de natuur is het gebruikelijk dat er soms hele groepen diersoorten worden verdreven door soorten die evolutionair beter bedeeld zijn. Bijvoorbeeld, tijdens het Eoceen werden grote delen van de wereld bevolkt door vroege primaten. Ze leken ietwat op apen, maar hadden een langere snuit die meer leek op die van een hond, en hadden kleinere hersenen. Echter verschillende opeenvolgende veranderingen zorgde ervoor dat sommige van die halfapen een meer typischere apekop kregen en een groter brein. Hiermee was de 'echte' aap ontstaan. Met de voordelen die met deze veranderingen gepaard gingen konden apen beter uit de voeten dan de halfapen, en deze begonnen dan ook de wereld over te nemen ten kost van die laatste. Al spoedig was de hele wereld overgenomen door moderne apen. Het kan echter gebeuren dat een kleine populatie door natuurlijke barrires wordt gesoleerd van de rest van de wereld, waardoor de nieuwkomers niet kunnen concureren met de oude levensvormen en deze dus ook niet kunnen uitroeien. Op die manier ontstaan levende fossielen. In het geval van de halfapen was er een populatie op Madagascar de dans ontsprongen, hun eiland was intussen weggedreven van het vasteland. Doordat er nooit ware apen op Madagascar zijn gekomen leven hier nog steeds primitieve primaten, met een hondachtige snuit en kleine hersenen.
Een gemeenschappelijk kenmerk van levende fossielen is zeldzaamheid, en isolatie. Zoals reeds vermeld leeft de Tuatara enkel nog in Nieuw-Zeeland en ook de Coelacanth is erg zeldzaam, niet voor niets duurde het zo lang voor men hem te pakken had. Degenkrabben zijn relatief minder zeldzaam, maar wel vergeleken met de mate waarin ze vroeger voorkwamen. Levende fossielen leven vaak gesoleerd, op een eiland, of diep in zee, overal slechts in kleine gebieden in kleine populaties. Door zich te vestigen in een gebied dat door natuurlijke grenzen wordt afgescheiden van de rest van de wereld stappen ze als het ware uit de strijd om het bestaan, ze kunnen niet meer worden verdreven door soortgenoten met betere mutaties, en blijven zodoende ten minste voor een gedeelte hun oorspronkelijke eigenschappen behouden. Dit scenario zien we overal in de natuur terug, ik gaf al de voorbeelden van de Tuatara en de halfapen die op eilanden zijn gesoleerd van de rest van de wereld , en andere voorbeelden zijn Australi, waar basale zoogdieren zoals Buideldieren en Monotremen hebben kunnen overleven doordat er geen placentaire zoogdieren van het vasteland in Australi konden komen (Dit gebeurde pas toen de mens arriveerde en honden en varkens meebracht, hierdoor zijn wilde honden ondertussen bezig de inheemse fauna uit te roeien, zo heeft de buidelwolf het al bijna tegen hen afgelegt.), of Zuid-Amerika, dat voor minstens 70 miljoen jaar gesoleerd is geweest van de rest van de wereld, ook hier leefde lange tijd Buideldieren en basale placentaire dieren zoals de Xenarthra.
Dit komt trouwens niet zomaar uit de lucht vallen, Darwin had dit in 1859 ook al door, hij schreef er over:
Met behulp van deze inzichten kunnen wij misschien enkele feiten begrijpen die opnieuw ter sprake zullen komen in ons hoofdstuk over geografische spreiding; bijv. dat de producties van het kleinere continent Australi vroeger zijn verdrongen, en klaarblijkelijk nog steeds verdrongen worden, door die van het grotere Europees-Aziatische gebied. Zo zijn vastelandproducties ook overal in hogere mate genaturaliseerd geraakt op eilanden. Op een klein eiland zal de wedloop om het leven minder hevig zijn geweest, en er zal minder modificatie en minder uitroeiing hebben plaatsgevonden. Daardoor, misschien, komt het dat de flora van Madeira volgens Oswald Heer lijkt op de uitgestorven tertiaire flora van Europa. Alle zoetwaterbekkens bij elkaar genomen beslaan een kleine oppervlakte, vergeleken met die van de zee of van het land; en dientengevolge zal de competitie tussen zoetwaterproducties minder hevig zijn geweest dan elders; nieuwe vormen zullen trager zijn gevormd, en oude vormen trager uitgeroeid. Het is ook in zoet water dat we zeven geslachten van glansschubbige vissen vinden, overlevenden van een orde die ooit overmachtig was; en het is in zoet water dat wij enkele van de meest abnormale vormen vinden die nu op aarde bekend zijn, zoals de Ornithorhynchus en de Lepidosiren die, net als fossielen, tot op zekere hoogte de verbinding maken tussen bepaalde ordes die thans ver van elkaar zijn verwijderd op de ladder der natuur. Deze abnormale vormen kunnen bijna levende fossielen worden genoemd; ze zijn tot op de dag van vandaag blijven bestaan omdat ze een beperkt gebied hebben bewoond en omdat ze derhalve aan minder hevige concurrentie blootgesteld waren.
Origin of Species; bladzijde 106/7
Bronnen:
1 The Marshall illustrated encyclopedia of dinosaurs & prehistoric animals, Cox & Palmer 1988.
Een levend fossiel is een soort die morfologisch zeer veel op zijn fossiele voorvaderen LIJKT : Het is een afstammingslijn die een lange periode van "stasis" heeft ondergaan .
Een beroemd voorbeeld is de coelacanth - Er zijn fossielen van deze groep van meer dan 300 miljoen jaar terug en die zeer op de moderne soorten LIJKEN . (= wat dus NIET wil zeggen dat ze identiek zijn aan fossiele soorten , noch dat de bepaalde extante soort ook daadwerkelijk fossiel is gedetermineerd /gevonden /ontdekt ...bovendien blijken de huidige soorten hoog gespecialiseerd en hoogstwaarschijnlijk aangepast aan andere habitats dan hun vermeende "identieke fossiele dubbelgangers )
Er zijn vroeger al twee werkhypotheses opgesteld die het mechanisme en de processen achter het verschijnsel, trachten te verklaren , maar er is (nog )geen algemeen geaccepteerde theorie .
Twee mogelijke redenen voor het bestaan van levende fossielen:
1.- De stabiliserende(" zuiverende" ) selectie (1) Sommige fossiele soorten leefden in vrij geïsoleerd habitat, zonder te veel of duidelijke concurrenten; er zal weinig selektiedruk geweest zijn zodat de omgeving voor die organismen redelijk stabiel is gebleven
Een moderne(?) variant van deze hypothese ziet de "aanpassende" evolutie binnen een bepaald geologisch stabiel en gegeven habitat als een proces van optimalisering ...wanneer de perfekte balans is bereikt zullen afwijkingen van die ontwikkelde norm worden weggezuiverd omdat ze minder fitness opleveren aan hun dragers
Veranderd echter het millieu ( klimaat/geologische gebeurtenissen/ veranderingen in de fysico -chemische samenstellingen )dan kan de evolutie (eventueel) opnieuw starten ....Sommige soorten zullen beter reageren op bijvoorbeeld klimaatsveranderingen(=standvogels ) dan andere ( bijvoorbeeld te laat terugkerende trekvogels )Uiteindelijk vallen er dan niches leeg die door de overlevers zullen worden ingenomen omdat ze minder concurentie krijgen in die niches van optimaal daaraan aangepasten die ondertussen zijn weggevallen ...
2.- Het Ontbreken van genetische variatie * Er is echter GEEN bewijsmateriaal dat levende fossielen genetische systemen bezitten die ( alle ) evolutieve verandering zouden kunnen verhinderen. = Ook de hoeveelheid eiwitpolymorfisme ( variatie in genprodukten en dus indicatoren van mutaties ) in extantemoderne en " levende fossielen" is gemeten dmv electroforese en bleek niet merkbaar verschillend (= bij de levende fossielen lager) tussen beide
levende fossielen zijn interessant om de volgende redenen:
1.- De moleculaire evolutie van levende fossielen verstrekt een opvallend voorbeeld van de afhankelijkheid tussen moleculaire en morfologische evolutie. 2.- Levende fossielen (zoals de longvis )verstrekken waardevolle informatie over de prijs van evolutieve veranderingen ---> overspecialisatie( optimalisatie )veroorzaakt ( misschien ) een vernauwing in de aanpassingsmogelijkheden ( of de onmogelijkheid bepaalde aanpassingen te ontwikkelen ) bij snelle veranderingen van het millieu 3.- Levende fossielen zijn een genetisch en morfologisch venster op volledig uitgestorven groepen ( Bijvoorbeeld de fossiele reptielen-orde der Ryncocephalia )
(1) Meestal stabiliseert de natuurlijke selectie. bv ..... Zo kan de gemiddelde lichaamsgrootte van een populatie , een hogere fitness bezitten dan extremen . De natuurlijke selectie handelt in dit geval tegen verandering in de lichaamsgrote, en houdt vormvan de leden van bevolking door tijd vrij constant. ( ten minste zolang het leefmillieu ( en zelfs de sociale interacties ) redelijk constant blijven )
De natuurlijke selectie kan ook richting geven én vernietigend zijn.
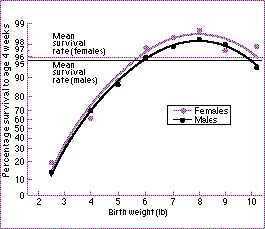
Een nog beter uitgewerkt voorbeeld is de stabiliserende selectie die het menselijk geboortegewicht binnen nauwe grenzen houdt ....
Borelingen die 8 pond wegen bij geboorte hebben een hoger overlevingskans dan zwaardere of lichtere babies . De grafiek is gebaseerd op neo-nativen geboren in Londen vanaf 1935 tot 1946. Van Karn en Penrose (1951).
The last European fossil occurrence of a coelacanth is from the Mid-Cretaceous of the English Chalk (Turonian, 90 million yearsago).
Here, we report the discovery of a coelacanth from Late Cretaceous non-marine rocks in southern France.
It consists of a left angular bone showing structures that imply close phylogenetic affinities with some extinctMawsoniidae.
The closest relatives are otherwise known from Cretaceous continental deposits of southern continents and suggest that the dispersal of freshwater organisms from Africa to Europe occurred in the Late Cretaceous.
Since Agassiz (1839) described a tail of Coelacanthus from the Permian of England, coelacanth fishes have long been known as fossils. They were thought to have become extinct some 75million years ago. The living coelacanth, Latimeria, from the western Indian Ocean, was described bySmith (1939), as recently was another from the Celebes Sea (Erdmann et al. 1998). Marine representatives of the Latimeria lineage are known from the Mid-Cretaceous (AlbianTuronian) chalk of England and Czech Republic with Macropoma (Forey 1998), and from the Late Cretaceous of North America with Megalocoelacanthus (Schwimmer et al. 1994).
Alongside the Latimeria lineage, freshwater mawsoniid coelacanths occurred in the Late Cretaceous of Africa and Madagascar as well as in the Early Cretaceous brackish/freshwater deposits of South America.
Here, we provide the first direct evidence, to our knowledge, of a continental coelacanth in the European Late Cretaceous in the form of an angular referable to a mawsoniid coelacanth. Geologically, the new find from France is about 20million years younger than Macropoma. The specimen shows affinities with the MawsoniaAxelrodichthys complex and indicates brackish or, more likely, freshwater dispersals between Africa and the European archipelago during the Late Cretaceous.
FOSSIL MATERIAL
The fossil coelacanth described here was found in 2004 during excavations at the Early Maastrichtian locality of Cruzy (southern France). The vertebrate remains were found in a lens of variegated clays cut by unfossiliferous sandy channels. The sediments of the Cruzy locality were deposited in continental environments, probably a floodplain. The fossil vertebrate assemblage comprises isolated remains of gars, lissamphibians, squamates, turtles, crocodiles, pterosaurs, ornithopod, sauropod and theropod dinosaurs, enantiornithine birds and mammals (Buffetaut et al. 1999).
RESULTS
Our identification of the specimen as a coelacanth angular is based on the rectangular outline shape, the pattern of the mandibular sensory canal and the openings from that canal (figure 1a, m.s.c.), the path of the groove for the external mandibular ramus of the facial nerve (figure 1a, gr.VII.m.ext) and the presence of a ridge on the medial side which marks the floor of the adductor fossa (figure 1a, r). The specimen is referred with confidence to the MawsoniaAxelrodichthys complex because of an ornamentation consisting of coarse ridges radiating from a point in the posterior third of the bone, an inflated lateral surface, the few slit-like ventral openings of the sensory canal (figure 1b, arrows) and the well-marked medial contact surface with the prearticular (figure 1a, con.Part). Coelacanths other than Mawsonia, Axelrodichthys, Lualubaea and Megalocoelacanthus show an ornament upon the angular that consists of enamel-topped tubercles or ridges, or is absent altogether; a mandibular sensory canal that opens through rounded pores; an ill-defined area of contact between the angular and prearticular, and a centre of ossification on the angular that is close to the deepest part of the bone (Forey 1998). Among these coelacanths our specimen may be distinguished from Megalocoelacanthus because that taxon shows an angular with a posteriorly directed opening for a special subopercular branch of the sensory canal, which indicates affinities with the latimeriids (Forey 1998). We are less certain about its distinction from other coelacanths within the MawsoniaAxelrodichthys complex: for this we would need more material.
Left angular of a mawsoniid coelacanth from the Early Maastrichtian locality of Cruzy, southern France (M2205, collection Association culturelle, archéologique et paléontologique de l'Ouest biterrois). (a) Internal views, (b) lateral views (more ...)
DISCUSSION
Late Cretaceous coelacanths are represented only by the marine latimeriids and the fresh water mawsoniids. Mawsoniid coelacanths diverged from the latimeriids in the Early Mesozoic. Mawsoniid coelacanths were exclusively Mesozoic and most lived in fresh water. Typical members, such as Mawsonia and Axelrodichthys, were heavy-bodied and showed an unusual pattern of fusion between bones in the skull roof. Some grew to a large size, estimated at over 2m, and rivalled the size of Latimeria today (Wenz 1981).
Late Cretaceous occurrences of Africa and Madagascar (Gottfried et al. 2004) are exclusively from fresh water. Both Mawsonia and Axelrodichthys are known in the Early and Middle Cretaceous of both sides of the proto South Atlantic (Cavin & Forey 2004) showing a vicariant biogeographic pattern (Maisey 2000). The exclusive occurrences of mawsoniids in fresh waters of Africa (Cavin & Forey 2004) and Madagascar (Gottfried et al. 2004) during the Late Cretaceous strongly suggest that mawsoniids reached the European archipelago by dispersal from Africa. We cannot exclude the possibility that dispersal polarity was from Europe to Africa. However, in the absence of contradictory information, we accept that the earlier and more widespread occurrence on and around the southern continents suggests that this is the more likely starting point. Although reconstructed maps show a wide marine barrier between Africa and the European archipelago in the Late Cretaceous (Hay et al. 1999; Scotese 2003), dispersal events have already been detected for tetrapods (Buffetaut 1989; Le Loeuff 1991) and freshwater fishes (Cavin et al. 1996), probably via a discontinuous route (Rage 1997).
Acknowledgments
We thank all those who took part in the excavations and in the preparation of specimens, in particular the members of the Association culturelle, archéologique et paléontologique de l'Ouest biterrois (ACAP). The excavation works were supported by the ECLIPSE Programme of the Centre National de la Recherche Scientifique (CNRS) and the ACAP. L.C.s research was supported by a Marie Curie Individual Fellowship funded by the Swiss Federal Office for Education and Science (grant no. 02.0335).
REFERENCES
Agassiz, L. Recherches sur les poissons fossiles. vol. 2. Published by the author, Petitpierre; Neuchâtel: 1839.
Buffetaut E. Archosaurian reptiles with Gondwanan affinities in the Upper Cretaceous of Europe. Terra Nova. 1989;1:6974.
Buffetaut E, Le Loeuff J, Tong H, Duffaud S, Cavin L, Garcia G, Ward D, L'association Culturelle, Archéologique Et Paléontologique De Cruzy. Un nouveau gisement de vertébrés du Crétacé supérieur à Cruzy (Hérault, Sud de la France). C. R. Acad. Sci. Paris, Sciences de la terre et des planètes. 1999;328:203208.
Cavin, L.; Forey, P.L. New mawsoniid coelacanth (Sarcopterygii: Actinistia) remains from the Cretaceous of the Kem Kem beds, SE Morocco. In: Tintori A, Arratia G. , editors. Mesozoic fishes III. Dr Pfeil Verlag; Munich: 2004. pp. 493506.
Cavin L, Martin M, Valentin X. Découverte d'Atractosteus africanus (Actinopterygii: Lepisosteidae) dans le Campanien inférieur de Ventabren (Bouches-du-Rhône, France). Implications paléobiogéographiques. Revue de Paléobiologie. 1996;15:17.
Erdmann M.V, Caldwell R.L, Kasim Moosa M.K. Indonesian king of the sea discovered. Nature. 1998;395:335.
Forey, P.L. Chapman & Hall; London: 1998. History of the coelacanth fishes.
Gottfried, M.D.;Rogers, R.R.; Curry Rogers, K. First record of Late Cretaceous coelacanths from Madagascar. In: Arratia G, Wilson M.V.H, Cloutier R. , editors. Recent advances in the origin and early radiation of vertebrates. Dr Pfeil Verlag; Munich: 2004. pp. 687691.
Hay, W.W., et al. An alternative global Cretaceous paleogeography. In: Barrera E, Johnson C.C. , editors. Cretaceous ocean/climate systems. Geological Society of America; Boulder, Co: 1999. pp. 148.
Le Loeuff J. The CampanoMaastrichtian vertebrate faunas from southern Europe and their relationships with other faunas in the world; palaeobiogeographical implications. Cretaceous Res. 1991;12:93114.
Maisey J.G. Continental break up and the distribution of fishes of Western Gondwana during the Early Cretaceous. Cretaceous Res. 2000;2000:281314.
Schwimmer D.R, Stewart J.D, Williams G.D. Giant fossil coelacanth of the Late Cretaceous in the eastern United States. Geology. 1994;22:503506.
Smith J.L.B. A living fish of Mesozoic type. Nature. 1939;143:455456.
Wenz S. Un coelacanthe géant, Mawsonia lavocati Tabaste, de l'Albien-base du Cénomanien du sud marocain. Annales de Paléontologie (Vertébrés). 1981;67:120.
21-03-2008 om 15:58
geschreven door Tsjok45
Meer weerleggingen (1)
Meer weerleggingen van creationisten over levende fossielen
YECs weten totaal niet wat een "levend fossiel" in werkelijkheid is ?:
In elk geval blijken hun aangevoerde voorbeelden steeds weer , niets meer te zijn dan : of fossiele en extante dieren /organismen van verschillende soorten, die door hen worden beschouwd( of ingedeeld volgens hun nep-wetenschap) als behorend tot dezelfde geschapen "kind "( = Baramin , Baramoon , MPG ... etc) of kompleet verkeerde identificatie uit onbegrip , onkunde en onbenul ... of volgehouden vervalsingen en leugens
Volgens YECs, weerleggen de "levende fossielen" zowel, het feit als de theorie van de evolutie (gretig ovrgenomendoor "andere " creationisten-scholen en de oude aarde, terwijl ze onwetend zijn ( of zich zo voordoen ) van het feit dat er geen geschreven biologen -"reglement " en/of natuurlijke evolutiewetten bestaan , die eisen dat de moderne soorten altijd moeten verschillen van fossiele soorten
Ze(willen niet )begrijpen ook nooit dat de evolutie met verschillende snelheden kan werken / zelfs binnen dezelfde stamlijn ....
Hetzelfde dingetje wordt ook beweerd over "coelacanthen "; die " schepselen " zijn NIET veranderd ....maw ze proberen deze voorhistorische vissen te gebruiken om de evolutie te weerleggen door te beweren dat het vinden van levende coelacanthen (identiek aan de oude) de evolutietheorie en de oude aarde weerlegt : aangezien deze vondsten toch tegenspreken dat die vissen 70 miljoen jaar geleden zijn uitgestorven , toch ? ... terwijl hun nu gevonden huidige afstamelingen nog steeds hetzelfde zijn .... Creationisten blijven wel onwetend ( of hun toehoorderd worden onwetend gehouden ) van het feit dat de levende coelacanthen , ( zowel deComoren als de sulawesi Latimeria )/totaal verschillende soorten zijn dan de uitgestorven soorten Coelacanthidae
Het is duidelijke dat het "levende fossiel" van al de creationisten niets meer is dan een extante (in leven zijnde ) soort die in werkelijkheid verschilt van (gevonden ) fossiele soorten --> ; waarvan er duidelijk veel kunnen zijn die hoogstwaarschijnlijk verwant zijn--> ipv een tegenbewijs is het zelfseen ondersteuning van de gemeenschappelijke afstamming ( de stamboom van het levende ) van de beschouwde soorten en het fossiele archief ....
" De Coelacanth, werd lang beschouwd als een reeds -70 MY uitgestorven voorbeeld van een vis-tetrapod overgang (transitionnal), maar werd levend en wel vandaag gevonden en onveranderd van vorm "... Bron: Morris, Henry M., 1974,
Antwoord :
1.- Met de moderne coelacanth werd (in 74 ) de soort chalumnae Latimeria, (in de familie Latimeriidae) bedoeld .
* Fossiele "coelacanthen" zijn afkomstig uit andere genera / families van de Coelacanthidae, en zijn beduidend verschillend in zoverre dat zij kleiner zijn en niet berschikken over bepaalde interne structuren. * Noch zijn het diepzeevissen ( =de huidige twee soorten zijn dat wél ;ze sterven door decompressie wanneer ze worden bovengehaald ) * Het genus Latimeria heeft tot nu toe geen enkele fossiele vertegenwoordigers , daarom kan het geen " levend fossiel" kan zijn ( in de zin die de creationisten daaraan geven ) * De diepzee is trouwens een zeer slechte plaats omuberhaupt fossilisatie toe te laten
2.- Maar zelfs als moderne coelacanthen en fossiele "coelacanthen " van dezelfde species( = alleen maar onderhevig aan zogenaade microevolutie ) waren: dan nog zou er geen ernstig probleem voor de evolutie zijn. * De evolutietheorie zegt niet dat alle organismen moeten evolueren( of aan de dezelfde snelheid moeten blijven evolueren ) . * In een ( relatief ) onveranderlijk milieu,waaraan een species in de loop der tijd optimaal is aangepast zou de natuurlijke selectie neigen om dingen grotendeels morfologisch onveranderd te houden( = purifying evolution ) .
3.-Coelacanthen bezitten veel primitieve eigenschappen in vergelijking met de meeste andere vissen, * Eens waren ze de enig bekende specimens die iets te maken hadden met de vis-tetrapoda -overgang. Ondertussen zijn verscheidene andere fossielen ontdekt die de vis-tetrapod overgang vrij goed aantonen.
Lees Forey, Peter L., 1998. History of the Coelacanth Fishes. London: Chapman & Hall.
Alhoewel het eigenlijk niet nodig is ...wil ik hier ook eventjes in gaan op de YEC- claim dat de "levende fossielen " een oude aarde weerlegt ( let er op dat deze weerlegging afkomstig is van de oude-aarde creationisten (OEC) wat dezaak alleen maar hilarisch maakt )
Bepaling van de geologische levensduur van een soort ( of een hogere classificatie-categorie )
Wanneer een fossiel in een afzetting wordt ontdekt , zoeken de paleontologen het geologisch "oudste" en het geologisch "jongste" specimen op; om te bepalen hoe lang de vertikale verspreiding van die soort in de geologische kolom aanwezig is ...
Bijvoorbeeld, als het oudste specimen van een bepaalde soort dinosaurus gevonden wordt in een rotslaag uit het Berrasian van het vroege -Krijt , dan bestond deze dinosaurus op zijn vroegst, ongeveer 140 miljoen jaar geleden (-140 Ma / soms ook -140 My ) . Als het jongste voorbeeld van deze zelfde soort dateert uit het Turonian van het meest-Recente Krijt( boven-krijt ) , dan stierf het, samen met zijn soort , uit ongeveer 90 miljoen jaar geleden.
Aldus, wordt voor die dinosaurus-soort een levensduur vastgesteld van -140Ma tot 90 Ma . Nadat deze data werden gepubliceerd, wordt diezelfde dinosoort opgegraven in een rotsbed daterend uit het Santonian van het boven-Krijt. Betekent dit dat de vroegere data... verkeerde gissingen waren? Neen , de data waren slechts onvolledig
Immers , nu loopt de geologische levensduur van die soort dino van 140 tot 84 Ma. Wetenschappers maken dit soort aanpassingen en up-dates voortdurend ; het is essentieel in het vergaren van de wetenschappelijke kennis . De nieuwe fossielen worden gevonden, breiden de bekende levensduur( en andere eigenschappen ) van het bestudeerde organisme uit met de nieuwe fossiele ontdekkingen. Het is geen truc of oplichterij namens de wetenschapper(s)... ze herinterpreteren slechts het beschikbare ( nu uitgebreidere )materiaalen en evidenties om toteen nog waardevoller conclusie te komen
Wat gebeurt er wanneer een fossiele heterogen groep , zoals coelacanth-achtigen , plotstwordt uitgebreid met levende exemplaren ? De geologische levensduur van dit type organismen was al bekend ... dat wordt dan uitgebreid!
Het betekent niet dat de aarde is jong, het levert zelfs geen goed bewijs voor een jonge aarde. Het geologische voorkomen van de coelacanth, werd uitgebreid van 400ma t/m -65 miljoen jaar geleden tot 400 Ma t/m heden. Verder kan er niets uit dit gegeven worden afgeleid .
Moest een enigzins op onze Dino lijkend wezens , vandaag ergens "levend " worden gevonden ( het monster van loch ness bijvoorbeeld ) dan zouden de YEC's , die vondst natuurlijk claimen als een "levend fossiel"....Maar voor de echte wetenschappelijke wereld zou het slechts gaan over de verlenging van de levensduur :van 140 Ma t/m het heden . Er zou helemaal geen paniek in de wetenschappelijke wereld uitbreken ... en men zou daar ook niet benieuwd zijn of de aarde werkelijk slechts 6.000 jaar oud is ____ De rest van het reeds gevestigde wetenschappelijk bewijsmateriaal in de natuurwetenschappen ,wijst trouwens duidelijk op een oude aarde _____
De aanwezigheid een het levend fossiel is op geen enkele manier bewijsmateriaal voor een jonge aarde.
Ja, er zijn groepen van fossielen die allang mee-draaien, maar er zijn er nog miljoenen anderen die lang geleden zijn uitgestorven en er is geen bewijsmateriaal dat zij vandaag nog ergens in leven zijn.
Daarom hebben wij zowel miljoenen uitgestorven soorten, en een handvol (waarschijnlijk honderden) zogenaamde "levende fossielen" die een hiaat van miljoenen jaren in het fossielen archief , in hun geschiedenis bezitten . Het bewijsmateriaal steunt miljoenen jaren van uitstervingen , en een oude aarde.
Maar elk " levend fossiel" dat wordt ontdekt zal elke YEC als bewijs van een jonge aarde aanvoeren .
Is ID de wetenschappelijke (sic) uitleg voor creationiisten 150jaar Darwinisme... De evolutietheorie gewikt en gewogen David sorensen en revolutietheorie lalala DE VERRIJZENIS VAN DE JONGE VERDRONKEN KOE » Reageer (21) ID TOVERDOOS OVER SLECHTE ID-EETJES ,RESISTENTIE & TOVERDOCTORS c) OEC d) YEC
Een oeroud spoor Denisova : ZUID SIBERIË DENISOVA - mens
DMANISI AAP OF MENS DE OUDSTE ? » Reageer (1)Nakalipithecus nakayamai (<) Alweer eentje ? ..... De tand des tijds / Heidelberg-mens DE HEUPEN VAN EVA CASABLANCA MAN /erectus Hobbit is aparte soort ? » Reageer (2) NOG EEN BENDE BIJTERS POLONAISE met neanderthaler TANDEN UIT DE QESEM GROT Neanderthaler genoom Mitochondriale Genenkaart van Neanderthaler Xuchang mens
b)Biologie 1.-(EVOLUTIE ) NAS / IM Document 2008 (2) NAS / IM Document 2008 ( 1)
Cambrium & precambrium AVALON GABONESE chips
Evolutie in actie Opmerkelijk snelle adaptaties bij kroatische ruine-hagedissen:
EEN PLUIM VOOR CHINA Geef eens een pootje Pluimgewicht KLEIN DUIMPJE & DE REUS &VEREN Opnieuw gevederde Maniraptor Similicaudipteryx. Aerosteon riocoloradensis
Krokodillen
PAKASUCHUS KAPILIMAI Prestosuchus chiniquensis Terug naar zee viseter
Eritherium azzouzorum LJOEBA Darwinius masillae VLIEGENDE KATTEN ? Vleermuizen ALWEER EEN BELANGRIJK STUK UIT DE LEGPUZZEL Zee-zoogdieren ; Van de wal in de visgronden Indohyus / walvisevolutie Zeehonden-evolutie
*Blauwe en groene ( =nederlandse)teksten zijn meestal aanklikbare links *Engelse Wikipedia teksten verwijzen in de linkerkolom naar verschillende niet-engelse versies van het wikiartikel *Blauwe teksten tusssen "" , zijn voornamelijk ( gedeeltelijke)citaten afkomstig van mensen met andere meningen
Evolutie / Charles Darwin aan de basis: SELECTIE NATUURLIJK DE LEVENSBOOM UITSTERVEN DEEP TIME Biogeografie SEKSUELE SELECTIE Al jaren lang samen CO-EVOLUTIE De economie van de natuur Geleidelijke veranderingen
Unintelligent design:
IP Theorie Ons gebit , De appendix, Geboorte, De weg van het zaad , Nervus Laryngeus Recurrens, DNA , 75 mistakes , Het boek
Unintelligent design (1): Rechtop lopen Tomaso over het oog , oogspieren , Unintelligent Design (5): ZWEETVOETEN Unintelligent design (6): Vitamine C Unintelligent design (8): Kuitspieren









 million years ago. The living coelacanth, Latimeria, from the western Indian Ocean, was described by
million years ago. The living coelacanth, Latimeria, from the western Indian Ocean, was described by





 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}
{kind=link}