Dit blog is een onderdeel van EVODISKU WAT IS DE BEDOELING EN WAAR STAAT DIT BLOG VOOR
****Wie meent dat alles inmiddels wel over de evolutietheorie gezegd is en dat de discussie gesloten kan worden, ziet over het hoofd dat de wetenschap niet stil staat.
***Wie meent dat inmiddels het creationisme definitief het pleit heeft verloren en dat de discussie gesloten kan worden , ziet over het hoofd dat het "creationisme" is geevolueerd ( en zal evolueren ) in nieuwere mimicrytische vormen( meme-complexen ) zoals bijvoorbeeld het ID(C)
***Dit blog is speciaal opgezet om de aktualiteit binnen het evolutie-creationisme debat te volgen en van kommentaren te voorzien ... waartoe de lezers zijn uitgenodigd bij te dragen ...
Let echter wél op het volgende :
"Je bent een rund als je hier met religie stunt " ....
04-03-2008
De niet-zo-missing links gallerij (1)
Geen Fossiele Tussenvormen?
( Fedor steeman )
"Een veelgehoorde kreet uit het kamp van de creationisten is dat fossiele tussenvormen die de schakels vormen tussen de huidige diergroepen nog nooit zijn gevonden. Dit is pertinent onjuist, Er zijn van die creationisten die beweren dat er geen fossielen gevonden zijn die de overgangen documenteren tussen de huidige bekende hogere diergroepen (klassen, orden, etc.). Sterker nog: zij beweren dat dergelijke overgangen gewoonweg onmogelijk zijn, omdat zij zich niet kunnen voorstellen hoe dat plaats zou moeten hebben gevonden! "
Om hun ongelijk te bewijzen staat er op deze pagina een begin van een overzicht ( er zullen nog verschillende vervolglijstjes en regelmatige up-dates worden geplaatst ) Onder elke geslachtnaam staan Links ( blauw ;Engels , groen nederlands )die verbinden met wikipedia en andere serieuze sites Van elk geslacht staat een kleine beschrijving plus afbeeldingen ( en een " internet" link naar google afbeeldingen ) De basisbronnen : voor dit alles zijn te vinden op http://www.daaromevolutie.net/default.asp?action=show&what=art&ID=67&topic=&segm=3 http://nadarwin.nl/gallerij.html anticreato Tussenvormen bestaan niet evodisku Acanthostega
Van water naar land (vis tot viervoeter) Tussenvormen tussen vissen en tetrapoden (landdieren):
Oerkwastvinnigen (Wetenschappelijke naam: Rhipidistia (onderorde binnen de Spiervinnigen, Sarcopterygii)zijn uitgestorven spiervinnigen die leefden gedurende het Devoon (ca. 410-360 miljoen jaar geleden). Ze hadden gespierde vinnen en een asymmetrische staart. Hun schedel bestond uit twee scharnierende helften. Ze leefden voornamelijk in zee. Oerkwastvinnigen, met name Eusthenopteron worden beschouwd als de rechtstreekse voorouders van de amfibieën en daarmee van alle gewervelde landdieren (naast amfibieën ook reptielen, vogels en zoogdieren).
Een typisch voorbeeld voor een spiervinnige vis zoals deze tijdens de Devoon-periode leefden. De vinnen van zulke vissen vertonen al meer overeenkomst met de poten van viervoetigen dan met de vinnen van straalvinnige vissen (bv. een steur.).
osteolepis macrolepidotus Coelacanth uit het Devoon (the Orkney Islands, Scotland)
Een spiervinnige uit het Devoon die beschikte over benige vinnen. Er zijn al dezelfde botelementen in te ontdekken die we normaal in viervoetigen vinden zoals het opperarmbeen, het spaakbeen en de ellepijp.
De Prins van Miguasha. 'Eusthenopteron fossiel ,:lengte 30 cm: ,gevonden in Miguasha, in formaties van het boven- devoon (is ooit afgebeeld op ,een Canadese postzegel ) Collections du Musée de Miguasha.
Vergelijking van de skeletten van de lobvinnige Eusthenopteron (boven ) en de vroege primitieve amfibie Ichthyostega (onder ). (Colbert and Morales, Evolution of the Vertebrates, 4th ed., (New York: John Wiley a& Sons, 1991), 69.)
Een vis die sterk leek op Eustenopteron, maar nog meer weg had van viervoetigen door grote overeenkomsten in de schedel. De meest aan de zogeheten tetrapoden verwante bekende fossiele vis is Panderichthys. Deze soort kan daarom informatie verschaffen over de evolutie van de vinnen naar poten. Over de evolutie van borstvin naar voorpoot (met alles wat daarmee te maken heeft, zoals de ontwikkeling van een schoudergordel) was op basis van fossiele vondsten al het een en ander bekend (de borstvinnen en schoudergordel van Panderichthys zijn typisch tussenvormen tussen die van vissen en tetrapoden), maar voor de ontwikkeling van de aarsvin naar de achterpoot gold dat niet, wegens gebrek aan fossiel materiaal. Analyse van een exemplaar van Panderichthys rhomolepis dat in 1972 werd ontdekt in Letland heeft daarin nu meer licht gebracht.Juist de overgang van aarsvinnen naar achterpoten is van groot belang voor het inzicht in de evolutie van de voortbeweging op poten, want alle huidige tetrapoden hebben als het ware achterpootaandrijving: de achterpoten verschaffen het overgrote deel van de kracht om te lopen of te rennen. Daarom werd tot nu toe gedacht dat dit ook bij de vroegste tetrapoden het geval was. Dat blijkt nu niet het geval te zijn.
De Tiktaalik was vermoedelijk de link tussen vissen en landdieren. Hij leefde in het Devoon, 375 miljoen jaar geleden. Zijn naam komt uit het Inuktitut (de taal van de Inuit van Groenland en Canada) en betekent grote vis in ondiep water. Duidelijk een vis vanwege de aanwezige visschubben en vinnen. De ledematen zijn precies zoals die van viervoeters met het enige verschil de afwezigheid van tenen. Eigenlijk was het dus een vis met poten, maar zonder tenen! Zijn schouders zijn niet verbonden met de schedel zoals in vissen, waardoor het een nek heeft gekregen. En het heeft ribben precies zoals tetrapoden die gebruikt konden worden om het lichaam te dragen en te helpen met ademhalen.
Het pectorale skelet van Tiktaalik is een "overgangsvorm"tussen vis en amfibie (zie ; Shubin et al over "the pectoral fin of Tiktaalik" waarvan een samenvatting is tevinden op http://afarensis.blogsome.com/2006/04/)
Tiktaalik
(artist reconstruction Ted Daeschler)
Internet Zie ook vooral deze reactie --> TIKTAALIK , weet je ? (Tsjok45) ( een korte compilatie van een discussie die Bart Klink over o.a. deze overgangsvorm heeft gevoerd met een creationist)
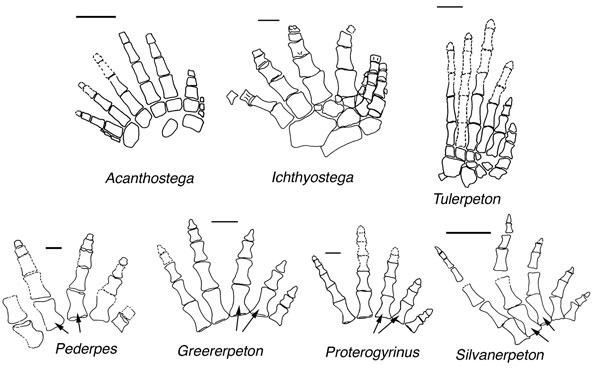
http://www.evolutionpages.com/pederpes%20finneyae.htm Reconstruction of pedes of various taxa. Pederpes, Greererpeton, Silvanerpeton and Proterogyrinus show asymmetrical metatarsals (see arrows) compared with those of the Devonian forms Acanthostega, Ichthyostega and Tulerpeton. In Acanthostega and Ichthyostega, the metatarsals are not clearly differentiated, and in Tulerpeton, if correctly interpreted, they are cylindrical but include some interarticulations. Scale bars, 10 mm. After reference (2)
a, Eusthenopteron (Late Devonian)3,21; b, Acanthostega (Late Devonian)12,14,16; c, Ichthyostega (Late Devonian); d, Pederpes (Early Carboniferous)7; e, Greererpeton (mid-Carboniferous)14,22; f, Pholiderpeton (Late Carboniferous)13,22. Skulls (grey) are in left lateral view, with hyomandibula/stapes (pink) in position. The left hyomanibula/stapes (not to scale) is shown in lateral (top left), dorsal (top right) and posterior (bottom) for each taxon (for Pederpes, lateral view is not available in detail). Acanthostega and Ichthyostega are shown in an unresolved polytomy with the Carboniferous forms.
a, GLAHMS 100815 Pederpes finneyae, photograph of holotype and only specimen. Scale bar, 100 mm; b, Map of the holotype, showing elements preserved on the reverse in grey. Scale bar, 10 mm. Fro, frontal; intemp, intertemporal; jug, jugal; lac, lacrimal; max, maxilla; nas, nasal; pmx, premaxilla; po, postorbital; pofr, postfrontal; prefr, prefrontal; preop, preopercular; qj, quadratojugal; scapcor, scapulocoracoid.
Dit was een zeer merkwaardig dier, het was een viervoeter, maar niet geschikt om over land te lopen, hiervoor waren de poten te zwak. Zwemmen kon het wel, hij beschikte over de zelfde zwemstaart als spiervinnige vissen en de spaakbeen-ellepijp verhouding was identiek aan die vanEusthenopteron. Als enige bekende viervoetige beschikte hij over 8 tenen per poot en over interne kieuwen. Waar we hier dus mee te maken hebben is een viervoetig dier dat in het water poten had ontwikkeld, maar verder was het nog vrijwel geheel een vis.
http://www.devoniantimes.org/Order/re-ichthyostega.html Een dier dat over zowel kenmerken van viervoetigen als spiervinnige vissen beschikte. Hij was in staat om met zijn voorpoten zich over het land voort te slepen, maar de achterpoten konden enkel als paddels fungeren.
Ichtyostega bezit een unieke mix eigenschappen met massieve schouders en zeven tenen , het bezat een amfibische leefstijl Acanthostaga is een tijdgenoot uit het laat devoon , die kompleetaquatisch was en warbij de ledematen fungeerden als peddels
De vondst van een gedeeltelijke onderkaak van Sinostega ,maakt deel uit van een err belangrijke sequentieele reeks van opeenvolgende ontdekkingen in china ; Psarolepis (1999), Achoania (2001), Styloichthys (2002) en Sinostega (2002)
incomplete linkse onderkaak ( 355 miljoen jaar oud/ laat-devoon )
a, Photograph (top) and interpretative drawing (bottom) of IVPP V13576, a lower-jaw ramus from the Ningxia Hui autonomous region, China. Note the absence of Meckelian ossification (Meckelian gap) between the prearticular and infradentaries (splenial and angular), and the narrow denticulated band along the top of the prearticular, both of which are diagnostic tetrapod features. Scale bar, 10 mm. b, Late Devonian palaeogeographical map (modified from ref. 7) showing the location of the new Devonian tetrapod (red dot) in relation to previously discovered specimens. Green numbering, Euramerican taxa; red, non-Euramerican: 1, Pennsylvania (Hynerpeton, Densignathus); 2, Scotland (Elginerpeton); 3, Greenland (Ichthyostega, Acanthostega); 4, Latvia (Obruchevichthys, Ventastega); 5, Russia (Tulerpeton); 6, New South Wales (Metaxygnathus). c, Map of modern-day China, showing the Ningxia Hui autonomous region in orange; green dot indicates the fossil site, close to the Yellow River. http://www.nature.com/nature/journal/v420/n6917/full/420760a.html
Een typisch voorbeeld voor een spiervinnige vis zoals deze tijdens de Devoon-periode leefden. De vinnen van zulke vissen vertonen al meer overeenkomst met de poten van viervoetigen dan met de vinnen van straalvinnige vissen (bv. een steur.).
Een spiervinnige uit het Devoon die beschikte over benige vinnen. Er zijn al de zelfde botelementen in te ontdekken die we normaal in viervoetigen vinden zoals het opperarmbeen, het spaakbeen en de ellepijp.
Panderichthys
Een vis die sterk leek op Eustenopteron, maar nog meer weg had van viervoetigen door grote overeenkomsten in de schedel.
Dit was een zeer merkwaardig dier, het was een viervoeter, maar niet geschikt om over land te lopen, hiervoor waren de poten te zwak. Zwemmen kon het wel, hij beschikte over de zelfde zwemstaart als spiervinnige vissen en de spaakbeen-ellepijp verhouding was identiek aan die van Eusthenopteron. Als enige bekende viervoetige beschikte hij over 8 tenen per poot en over interne kieuwen. Waar we hier dus mee te maken hebben is een viervoetig dier dat in het water poten had ontwikkeld, maar verder was het nog vrijwel geheel een vis.
Ichthyostega
Een dier dat over zowel kenmerken van viervoetigen als spiervinnige vissen beschikte. Hij was in staat om met zijn voorpoten zich over het land voort te slepen, maar de achterpoten konden enkel als paddels fungeren.
Tulerpeton
Een stevige schouder suggereerd dat het dier redelijk goed kon lopen. De voorpoot bevat 6 tenen, de achterpoot 7.
Het oudst bekende reptiel dat behoord tot de orde der Synapsiden. Een klein, hagedisachtig dier. Verassend genoeg beschikt het over enkele kenmerken van amfibien (Reptielen waren nog maar net ontstaan uit amfibien.).
Een verwant van Dimetrodon, een 3 meter lang, varaanachtig dier.
65%
Biarmosuchus
De oudst bekende Therapside, de voorgaande dieren waren Pelycosaurirs. Beide groepen vormen samen de orde der Synapsiden. Therapsiden staan bekend als de zoogdierachtige reptielen.
Een typisch voorbeeld van een zoogdierachtig reptiel, hij beschikt over kenmerken van zowel reptielen als zoogdieren. Waarschijnlijk nog wel koudbloedig.
65-75%
Pristerognathus
En van de eerste dieren waarvan het coronoide process (De uitstekende vleugels aan de achterkant van de onderkaak.) door het Synapside slaapvenster naar buiten steekt. Hierdoor wordt de basis gelegd door het Squamosale-Dentale kaakgewricht van zoogdieren. Ook bij alle volgende dieren is dit het geval.
Een Therapside wiens schedel en tanden sterk aan zoogdieren doen denken, maar waarvan de staart, heupen en kaakgewricht meer weg hebben van die van reptielen. De samenstelling van de onderkaak is een intermedium tussen reptielen en zoogdieren. Voor zover dat valt af te leiden zou Cynognathus misschien een vacht kunnen hebben gehad (onzeker).
Eveneens een Therapside wiens schedel en tanden sterk aan zoogdieren doen denken, hoewel de onderkaak nog niet helemaal die van een zoogdier is en de oogkas geen contact heeft met het Synapside slaapvenster. Het is het oudst bekende dier wiens ribben tot de orst zijn gereduceerd, wat warmbloedigheid suggereerd. Kleine putjes in de snuit geven de plaats van snorharen aan. Een zeer zoogdierachtig dier, maar net geen zoogdier.
85%
Probainognathus
Een zeer zoogdierachtig dier, maar de oogkas heeft nog net geen contact met het Synapside slaapvenster, en de onderkaak bestaat nog niet helemaal uit het Dentale been.
Een dier dat jarenlang voor een zoogdier is versleten, maar toen werd ontdekt de onderkaak nog niet helemaal uit het Dentale been bestond werd de ware identiteit achterhaald. Het skelet lijkt als twee druppels water op dat van eierleggende zoogdieren (monotremata).
Doordat de onderkaak geheel uit het Dentale been bestaat en de oogkas contact heeft gemaakt met het Synapside slaapvenster/achterhoofd wordt dit dier als een volledig zoogdier beschouwd. De kaakverbinding is uniek, het is een combinatie van het reptielachtige Quadrategewricht en het zoogdierachtige Squamosal-Dentale kaakgewricht.
100%
Steropodon
Het oudst bekende lid van de familie der Monotremata (eierleggende zoogdieren).
Dinosaurir uit de familie Dromaeosauridae, een zusterfamilie van de vogels. Door het polsbeentje was dit dier in staat om de zelfde vleugelslagen te maken als vogels. Sommige Dromaeosaurirs waren in het bezit van veren (Zoals Microraptor.).
Unenlagia
Een vogelachtige dinosaurir, die in staat was om zijn voorpoten op te vouwen en langs zijn lijf te houden zoals een vogel doet.
Dinosaurir met symmetrische veren aan zijn staart en armen. Symmetrisch betekend ongeschikt om te kunnen vliegen of zweven, toch was dit de eerste stap op weg naar vliegende vogels.
Een dinosaurir met asymmetrische veren en achterpoten die geschikt waren om het dier op een tak te laten zitten. Aan de andere kant vertoont het skelet ruim 20 reptielachtige kenmerken zoals de aanwezigheid van gastralia, tanden, een benige staart, een fibula en vleugelklauwen, en de afwezigheid van een tarsometatarsus, een carpometacarpus, een pygostyle, een borstbeen en vergroeide rugwervels.
Sinornis
Primitieve vogel, waarschijnlijk in staat om te vliegen. Vogelachtige kenmerken: een Pygostyle en korte staart, een borstbeen, en de voorpoten waren wellicht in staat om mee te kunnen vliegen. Reptielachtige kenmerken waren oa.: gastralia, tanden, geen snavel, reptielachtige heupen en ribbenkast, geen tarsometatarsus, en het kuit -en scheenbeen waren nog evenlang. Sinornis wordt beschouwd als de meest reptielachtige vogel, behalve Archaeopteryx.
Ongeveer het zelfde als Sinornis, maar iets moderner door een vrij moderne vogelhand en de afwezigheid van gastralia. Nietenim beschikte het dier nog over veel reptielachtige kenmerken zoals het ontbreken van een tarsometatarsus, onvergroeide rugwervels, een evenlang kuit -en scheenbeen en reptielachtige heupen en ribbenkast. Iberomesornis is gevonden in 1992 in Spanje.
Confuciusornis
Primitieve vogel, maar weer iets moderner door de aanwezigheid van een verhoornde snavel en geen tanden.
Een nogal grote vogel, 2 meter lang, uit het late Krijt. Hesperornis was niet in staat om te vliegen en waarschijnlijk ook niet om te lopen, maar was aangepast aan een leven in het water, zoals de hedendaagse Pinguin. I veel opzichten was het een moderne vogel, hoewel hij nog steeds beschikte over tanden.
In veel opzichten een moderne vogel, met uitzondering van de aanwezigheid van tanden. Ichtyornis beschikte over een groot borstbeen, vergroeide rugwervels en een tarsometarsus.
Een klein paardachtig bosdier uit het Eoceen, zo'n 45 miljoen jaar geleden. Hyracotherium was zo'n 20 centimeter hoog en had per voet 4 hoefjes van voren en 3 achter. Verder een 44-delig gebit met korte hoektanden en laaggekroonde kiezen, per kaakhelft 4 valse en 3 ware.
Orohippus
In grote mate gelijk aan Hyracotherium, maar iets groter, en de middelste teen droeg een beetje meer gewicht ten koste van de andere tenen.
Dit dier had een meer paardachtige lichaamsvorm dan de voorgaande dieren, het was zo'n 24 centimeter hoog en had nu drie tenen aan zowel de voor -als achterpoot.
Zo'n 60 centimeter hoog. Dit dier was te oordelen naar het gebit zowel een blad -als graseter. (De voorgaande dieren waren uitsluitend bladeters.) De kiezen waren hoger gekroond dan die van de voorgaande dieren.
Parahippus
Zo'n 90 centimeter hoog. Drietenig (Maar de twee buitenste tenen waren flink in omvang afgenomen.), een blad -en graseter. Kiezen nog hoger gekroond
Zo'n 90-100 centimeter hoog. Drietenig, maar de buitenste tenen raken niet de grond. Kiezen volledig hoog-gekroond, met een dikke laag cement. (Bij alle latere soorten blijft dit behouden.) Een graseter.
Een landzoogdier uit de familie der Mesonychiden, dit waren vleesetende hoefdieren.
Pakicetus
Dit dier leek sprekend op een Mesonychide, maar vertoont kenmerken van walvissen in het achterste van de schedel. In feite is het een op het land levende walvis.
Ambulocetus
De naam van dit dier betekent: "Lopende walvis die zwemt". Ambulocetus zag er uit als een 3 meter lange otter of een harige krokodil, maar vertoond duidelijke kenmerken van walvissen. Het vormde de eerste stap die de walvisfamilie zette op weg naar een leven als zeedier.
Rodhocetus
Rodhocetus was de verbeterde versie van Ambulocetus, hoewel hij nog beschikte over stevige achterpoten kon hij al goed zwemmen in open zee. (Dat weten we omdat de chemische samenstelling van de tanden onthulde dat het dier zout water dronk en bovendien werden de resten gevonden in gesteente die ooit zeebodem vormde.)
Dorudon
Een dier dat volledig aan het waterleven was aangepast, maar nog geen moderne walvis. Hij beschikte nog niet over een spuitgat, baleinen of echo-locatie. Vergelijk de schedel van Dorudon eens met die van Rodhocetus.
Squalodon
Een dier dat op weg was een moderne tandwalvis te worden. Het spuitgat was al redelijk gevormd, maar de driehoekige kiezen die al sinds Pakicetus aanwezig waren zitten er nog steeds in.
Van aap tot mens
Geslachtsnaam
Beschrijving
Afbeeldingen
Sahelanthropus
Oudst bekende rechtoplopende aap (8 miljoen jaar geleden.).
Ardipithecus
Rechtoplopende apesoort die ongeveer tussen Sahelanthropus en Australopithecus moet hebben geleefd. Alleen fragmentarisch materiaal bekend.
Het geslacht dat voorafging aan ons moderne geslacht Homo. Van Australopithecus zijn verschillende soorten bekend waaronder A.afarensis en A.africanus. Van die laatste zijn verschillende goed bewaarde schedels bekend en van afarensis een redelijk goed bewaard skelet (Lucy). Tevens talloze fragmetarische resten. Australopithecus zag er grotendeels uit als een aapachtige, maar liep op twee benen en vormt in sommige opzichten een intermedium tussen apen en mensen.
Soortnaam
Homo habilis
De oudst bekende soort van het geslacht Homo. Habilis had een meer plat, mensachtig gezicht dan Australopithecus. De hersenomvang bedroeg 47% (630 cc) van de omvang van een modern menselijk brein. In de buurt van habilisresten zijn de oudste werktuigen gevonden.
Homo rudolfensis
Een vroege mensachtige die ongeveer ter zelfder tijd leefde als habilis. Hij was en beetje groter dan die laatste, maar zeldzamer. Rudolfensis wordt in het algemeen niet geacht de directe voorouder van de huidige mens te zijn, hoewel sommige menen dat hij de nakomeling is van een in Afrika gevonden hominide: Kenyanthropus platyops en zelf de voorouder van de moderne mens. De meeste paleo-antropologen houden het echter op habilis als onze directe voorouder.
Samen met een verwante soort; Homo ergaster is Homo erectus een redelijk mensachtige soort. De vondst van een bijna kompleet skelet, dat bekend staat als de Turkana Boy, laat zien dat het lichaam vrijwel mensachtig is. De schedels vertonen daarentegen nog wat verschillen, de herseninhoud bedroeg 74% van een modern menselijk brein, de kaken en tanden waren nog wat groter en er bevonden zich nog beenwallen boven de ogen. Resten van erectus en ergaster zijn gevonden in Afrika en Azi tot diep in Indonesi. Stenen vuistbijlen worden in de buurt van hun resten gevonden.
Homo heidelbergensis
De gemeenschappelijke voorouder van de moderne mens en de Neanderthaler, de hersenomvang bedroeg 93% van een modern menselijk brein.
Homo neanderthalensis
De Neanderthaler, leefde tijdens de ijstijd in Europa en Azi. Geen voorouder van de huidige mens. Neanderthalers waren klein, maar zeer gespierd vergeleken met moderne mensen. Ze beschikte over een brein dat 12% groter is dan dat van moderne mensen.
Homo sapiens
Moderne mens, ontstond 150.000 jaar geleden.
29-05-2008 om 12:31
geschreven door tsjok45
27-05-2008
Tetrapoden ten voeten uit.
Tetrapoden ten voeten uit. Over vroege en moderne spiervinnige vissen en het ontstaan van de eerste tetrapoden.
Een artikel van Tim Spaan.
In 1837 kreeg de meest gerespecteerde anatoom en paleontoloog van Engeland, Richard Owen ('Sir' moest hij nog worden op dat moment.), in zijn laboratorium een geconserveerd exemplaar van een vis die was gevangen in Gambia in West-Afrika onder ogen. In zijn natuurlijke leefomgeving groef dit dier zich tijdens het droge seizoen in in de modder, en bleef daar maandenlang zitten in een hol van een halve meter diep. Het dier had een normaal visselijf, een zwemstaart en tamelijk lange, sliertachtige vinnen. Na deze oppervlakkige observatie ging Owen het exemplaar ontleden om meer te weten te komen over het innerlijk van de vis. In de bek bevonden zich de vreemdste gebitselementen die hij ooit gezien had, de tanden stonden vlak naast elkaar, als een grote geribbelde plaat. Twee van zulk soort platen waren aan het gehemelte bevestigd en werden gebruikt om prooien te vermorzelen. Owen had deze bizarre constructie eerder gezien, fossielen van zulk soort platen waren een tijdje geleden gevonden. Eerst aangezien voor delen van een schildpadschild, maar later herkend als gebitselementen. Met het intreden van de radiokoolstof dateringsmethoden zouden deze fossielen door geologen op 220 miljoen jaar oud worden gedateerd, maar ook Owen zag dat het fossielen van hoge leeftijd moesten zijn. Maar nu had hij het volledige dier, in moderne vorm met huid en vlees er aan. Hij sneed het visje open en onderzocht de ingewanden, spreidde de kieuwen aan hun benige bogen uit als een waaier. In de borstkas vond hij de zwemblaas. De meeste vissen beschikken hierover, en gebruiken deze met gas gevulde holte om hun drijfvermogen gelijk te stellen aan de zwaartekracht die hen naar de zeebodem trekt. Met zo'n zwemblaas hebben vissen dus geen angst om naar de zeebodem te zinken of aan het oppervlak te blijven drijven, een heel nuttige constructie dus. Owen onderzocht de zwemblaas van de vis en ontdekte hierin tot zijn stomme verbazing een paar lange zakken met een honingraatstructuur die met het hart verweven waren. Hij had geen andere keus dan vast te stellen dat het longen waren. Maar dit was toch te absurd voor woorden, een vis met longen??
Neoceratodus forsteri De Australische longvis (Owens exemplaar was een Afrikaanse Lepidosiren paradoxa.) Klik voor vergroting.
Hoewel het nog even zou duren voor Darwin de wereld op zijn kop zette met zijn revolutionaire evolutietheorie (Hij was nog maar net terug van zijn vierjarige wereldreis, en zou nog 25 jaar werken aan zijn theorie alvorens deze te openbaren.) waren er ook rond deze tijd al mensen die in Owens 'longvis' een overgangsvorm van vissen naar landdieren zouden willen zien. Het was ongeveer 20 jaar geleden dat de Franse bioloog Lamarck had geopperd dat verschillende dieren en diergroepen van elkaar zouden kunnen afstammen, en hij had al wat aanhangers gevonden. Vissen werden door deze vroege evolutionisten, als meest primitieve en basale gevervelden, beschouwd als de voorouders van de gewervelde landdieren. Op het eerste gezicht lijken de leefgebieden van land -en zeedieren een wereld van verschil. En dat een waterbewonend dier dat lucht zeeft uit het water door middel van kieuwen en zich voortbeweegt door een zwemstaart zich zou ontwikkelen tot een landbewondend dier dat lucht ademt door longen en zich voortbeweegt met ledematen leek praktisch onmogelijk. Maar toch zijn er in de natuur talloze voorbeelden hoe dieren de barrire tussen water land kunnen doorbreken. Zoetwatervissen, die leven in riviertjes en meertjes zijn al sowieso dichter met het land verbonden als echte zeedieren
Een slijkspringer, een amfibische vis en de bijna buitenaards lijkende -maar vriendelijk lachende- salamanderlarf Axolotl. Klik voor vergroting.
die ver buiten de kust leven. Waterrijk gebied als moerassen en wetlands zijn in feite kruispunten tussen land en water en hier is de natuurlijke barrire tussen deze twee elementen aanmerkelijk minder. In zulke gebieden leven dan ook dikwijls dieren die zowel in water als op land kunnen leven. Dieren als slijkspringers, vissen die zich met hun vinnen over land sneller kunnen voortbewegen dan mensen met hun benen en zelfs in bomen kunnen klimmen. Amfibien zoals kikkers, padden en salamanders, die als visje met zwemstaart en kieuwen ter wereld komen en terwijl ze opgroeien poten en longen ontwikkelen. Sommige amfibien krijgen zelfs nooit longen, zoals de Mexicaanse Axolotl. Deze salamander komt nooit verder dan zijn larvale stadium, en behoudt zijn rode externe kieuwen en visachtige zwemstaart. Dieren zoals dit doen de verschillen tussen land en zee vervagen, en brengen de gedachte dat landdieren zijn voortgekomen uit vissen binnen de grens van het voorstelbare. Maar Owen wees het idee van evolutie af, al besefte hij dat deze longvis angstvallig goed in het straatje van de Lamarckisten paste. Hij besloot verder te zoeken naar aanwijzingen die deze vis geheel van landdieren zou distantiren. Die vond hij, zo leek het, in de snuit. Bij alle landdieren staan de longen in verbinding met de neusgaten, zodat ze er mee kunnen ademen. Maar bij vissen zijn de neusgaten alleen openingen naar buiten toe, en ze dienen enkel om geursporen op te vangen. De geconserveerde longvis van Owen vertoonde geen sporen van een luchtpijp die naar de zwemblaaslongen leide. En hiermee besloot hij dat de longvis overgangsvorm -af was. Waarschijnlijk was Owens longvis niet goed genoeg geconserveerd, -of had een verstopte neus- want na verdere onderzoeken met nieuwe exemplaren werd ontdekt dat longvissen wl een inwendige neusgaten hebben, een soort neusgangachtige verbinding met de mondholte. Een andere anatoom, de Ierse Robert M'Donnel liet een levende longvis in een houten aquarium bij hem thuis bezorgen, alwaar hij het dier bestudeerde. Hij kwam tot de conclusie dat longvissen daadwerkelijk in staat waren om met hun zwemblaaslongen lucht in te ademen. Hierdoor overleven de dieren de lange periode dat ze in de grond zijn ingegraven. In het water maken ze evenwel gebruik van hun kieuwen om aan zuurstof te komen. Bij onderzoek naar de werking van het hart zag hij dat deze een sterke gelijkenis vertoond met die van een kikkervisje, en de longen zelf leken op die van een slang. Al met al had hij geen idee in welke orde hij dit dier moest indelen, het leek op een vis, maar toch ook weer op een amfibie. M'Donnel schreef: "We mogen aannemen dat Lepidosiren (De longvis) als gevolg hiervan een bijzonder belang krijgt, omdat hij, staande op de grens tussen twee hoofdafdelingen van de dierlijke schepping, op zichzelf een bewjs vormt van de vrijwel onmerkbare overgangen van de ene klasse naar de andere als we de grote ladder der natuur bestijgen. Ik ken geen dier dat er meer op berekend is je de theorie van Darwin te laten aanvaarden dan Lepidosiren.".
Tegenwoordig weten we dat de longvissen, die worden samengevat in de orde der Dipnoi (Letterlijk: "Tweelongigen".), een lange geschiedenis achter zich hebben. De oorsprong van deze dieren ligt in het Devoon, een tijdperk dat duurt van ongeveer 417 t/m 354 miljoen jaar geleden. En samen met bijvoorbeeld de familie van de Coelacanth behoren ze tot de orde der spiervinnigen (Sarcopterygii). Alle spiervinnigen bezitten -waarom zouden ze anders zo heten- gespierde vinnen met een aantal forse botten hierin. De vinnen van een longvis vertonen zelfs meer overeenkomsten met een menselijke arm dan met de vin van bijvoorbeeld een steur. Er zijn echter ook duidelijke verschillen, het schoudergewricht van tetrapoden bestaat uit een kop en kom, waarbij het uiteinde van het been de kop, en de holte in de schouder de kom vormt. Longvissen hebben hun schouders echter aan de kop vastzitten, en de kom aan het opperarmbeen. Desondanks worden longvissen nu beschouwd als de nauwste nog levende verwanten van tetrapoden (Nauwer dan bv. de Coelacanth of slijkspringers.).
De vin van Eusthenopteron.
Maar nog nauwere verwanten worden gevonden in het fossielenbestand. Rond 1880 vond een Canadese boer een prachtig ongeplet fossiel van een spiervinnige vis die Eusthenopteron werd genoemd, en in de jaren daarna zijn er nog meer gevonden die in goede staat verkeerden. Een levende Eusthenopteron zou er hebben uitgezien als een snoek, maar met de plompe kop en vlezige vinnen van een Coelacanth. Het zijn de vinnen die hem bijzonder maken, hierin bevinden zich botelementen die gewoonlijk alleen in de poten van landdieren voorkomen, zoals een humerus, radius & ulna in de voorvinnen en een femur, tibia & fibula in de achterste vinnen. Bovendien werd ontdekt dat de benige vinstralen in feite ongesegmenteerde vingers waren, dit houdt verband met de manier waarop vingers in een embryo worden gevormd:
Simpel gezegd ontstaan binnen een embryo de arm en hand uit een rechte staaf, de metapterygiale as, die van de onderarm in een rechte lijn doorloopt tot in de wijsvinger. De andere vingers zijn vertakkingen die aan het einde van de as ontspringen. Deze metapterygiale as is duidelijk terug te vinden in de vin van Eusthenopteron, en ook hier zijn de vinstralen vertakkingen van de as, net zoals de vingers van een tetrapode embryo. In dit licht kun je constateren dat de vinnen van Eusthenopteron ineengedrukte handen en voeten zijn. Of beter gezegd, onze handen en voeten uiteengetrokken vinnen zijn met gesegmenteerde vinstralen.
Hoewel dit een sterke aanwijzing is dat spiervinnige vissen als Eusthernopteron gestalte hebben gegeven aan de tetrapoden, was er begin 20e eeuw nog geen fossiel bewijs van de eerste echte poten zelf. "Tussen de oudste Amphibia en alle vissen zitten diepgaande structurele verschillen die nog niet door paleontologische vondsten zijn overbrugd", schreef een paleontoloog in 1915. In 1931 kwam hier verandering in. Een energieke 22-jarige Zweedse geoloog, Gunnar Sve-Sderberg, kreeg de leiding over een paleontologische expeditie naar Groenland. Hier waren al eens eerder skeletten en schubben van spiervinnigen uit het Devoon ontdekt, bij toeval, want de vinders behoorden eigenlijk tot een reddingsteam die een mislukte poolreiziger wilde redden. Gunnar Sve-Sderberg had genummerde labels bij zich om de gevonden fossielen te labelen. P-labels voor vissen en A-labels voor amfibien, heel optimistisch, want niemand had ooit nog een amfibie uit het Devoon gevonden. In de zomer van 1931 werkte de expeditie in de buurt van Gauss HalvØ aan de noordelijke en oostelijke helling van de Celciusberg. Er werd een behoorlijk aantal fossielen gevonden, waaronder 15 fragmenten van een platte schedel die zeker niet die van een vis was. Sve-Sderberg labelde ze met een A. Die herfst terug in Stockholm werden de resten steen tussen de botten verwijderd en werd de structuur ervan beter bestudeerd. Sve-Sderberg zag dat zijn eerdere conclusie juist was, hoewel een deel van de schedelbeenderen het patroon van die van spiervinnige vissen vertoonde had het een aantal kenmerken die enkel voorkomen bij tetrapoden. Sve-Sderberg noemde deze nieuwe evolutionaire schakel Ichthyostega, 'Ichtyo' (vis) naar zijn visachtige kenmerken, en 'Stega' (vlak) naar zijn platte, wc-bril vormige snuit. Hierna bezocht Sve-Sderberg elke zomer weer het fossielrijke gebied van Groenland, waarbij hij en zijn team grote hoeveelheden vissen en enkele nieuwe schedelfragmenten van Ichthyostega vonden. En fossiel gevonden in 1934 was zo mooi bewaard dat het op een fluwelen kussen werd vervoerd. Na vijf succesvolle jaren werd hij benoemd tot hoogleraar aan de universiteit te Uppsala, maar in datzelfde jaar bleek dat hij tbc had, en moest hiervoor zijn werk bijna geheel neerleggen. Hij stierf in 1948 in dezelfde zomer als waarin in Groenland schouders, poten en een staart van Ichthyostega werden gevonden. Toen bleek dat enkele fossielen die eerst voor vissenschubben werden aangezien in feite de ribben van Ichthyostega waren kon men alle gevonden delen samenvoegen en een reconstructie van het dier maken.
Ichthyostega (Behalve de tenen aan de voorpoten is het complete skelet gevonden.)
Degene die deze taak op zich nam was Erik Jarvik, de spiervinnigenexpert van de expeditie. De poten van Ichthyostega waren kort en plomp, maar bezaten de kenmerken van een tetrapode. In de schedel zag Jarvik een naad, op de plek waar spiervinnigen een gewricht hebben. De interne neusgaten werden van elkaar gescheiden van een dun laagje been, net als bij spiervinnigen, terwijl gangbare amfibien hier een zeer brede beenlaag hebben. Tussen de neusgaten werd ook een klein rostrolateraal botje gevonden dat enkel voorkomt bij spiervinnigen, en geen enkel ander amfibie. Maar het meest visachtige kenmerk was wel de staart. Tetrapoden hebben een betrekkelijk eenvoudige staart, een reeks taps toelopende wervels, ingebed in spierweefsel. De staart van vissen, en dan vooral een spiervinnige is een veel complexer geval, en dat mag ook wel, de staart is ten slotte de motor van het dier. Elke wervel heeft hier naar onder en boven lopende uitsteeksels. Hieraan zitten kleinere beentjes, de zogeheten radialen, en daaraan zit een brede waaier van vinstralen die, net als de schubben, uit een ander soort been bestaan. Door die ingewikkelde structuur kan een spiervinnige met de golvende bewegingen van zijn staart, een plotselinge spurt maken en weer even plots remmen. De staart van Ichthyostega is een kruising tussen die twee soorten staarten, de onderkant is die van een amfibie, terwijl de bovenkant identiek is aan een spiervinnigenstaart. De schedel vertoonde geen sporen van kieuwen maar wel resten van pre-opperculare botten (Platte J-vormige benige plaatjes in het kieuwskelet van vissen ter bescherming van de kieuwen.) op dezelfde plaats en positie als bij spiervinnigen. Deze en andere kenmerken maken Ichthyostega tot een kruising tussen spiervinnige vissen en amfibien. Het beest was verder een meter lang, had brede kaken en een gehemelte vol tanden, waarvan sommige op lange giftanden leken. De hele anatomie suggereerde dat het een roofdier was dat in zot water leefde en jaagde. We kunnen ons, dankzij de vondsten van paleontologen, een beeld vormen van hoe de wereld er tijdens het Devoon uitzag. Het land dat Ichthyostega aandeed was tijdens het Siluur gekoloniseerd door terrestrische planten. Planten waren hiermee de eerste levensvormen die aan land gingen, en in de tijd van Ichthyostega waren de eerste echte bossen gevormd, waarvan sommige bomen al hoogtes van 30 meter hadden bereikt. Ook waren er al dieren aan land gekomen, maar die hadden een meer bescheiden grote. De enige zeedieren die zich op dat moment hadden aangepast aan een leven op het land waren slakken en verder geleedpotigen, zoals vroege vormen van insecten, pissebedden, spinnen, schorpioenen en duizendpoten. Dit waren dus de enige dieren die in het Devonische woud leefden. Voor een in de tijd reizende mens zou het maar een vreemd gezicht zijn, een kompleet uitgestorven woud, met als enige dieren hier en daar een pissebed, die zich nog niet onder stenen verstopten bij een gebrek aan roofdieren. Een woud waar behalve het ruisen van de wind in de takken doodse stilte heerste, omdat zingende vogels nog niet bestonden. De temperatuur was in het Devoon tropisch, na een zeer koude periode in het Ordovicium.
Twee Placodermen uit het Devoon, Bothriolepsis (30 cm) en de veel grotere Dunkleosteus (3,5 tot 9 meter).
Maar waar het land tijdens het Devoon onderbevolkt was, waren de oceanen gevuld met een enorme diversiteit aan gewervelden die zijn weerga niet kent. Placodermen, pantservissen van soms 9 meter lang, met huiveringwekkende kaken waar ze elkaars pantsers mee trachten te kraken, vroege haaisoorten waarvan sommige een bizarre van tanden voorziene rugvin hadden, spiervinnigen waarvan sommige leken op de Coelacanth, en reusachtige longvissen. Ook Eusthenopteron was n van die dieren, en hoewel Ichthyostega een dier was dat met zijn poten enigszins op het land uit de weg kon was ook hij voor zijn voortbestaan afhankelijk van het water, wat zijn jachtgebied was.
Met de spiervinnige Eusthenopteron en de vroege tetrapode Ichthyostega was er al behoorlijk wat helderheid gekomen in de evolutie van de viervoetigen. Maar het plaatje was nog niet kompleet, aan beide zijden van Ichthyostega bevonden zich nog onoverbrugde gaps. De jaren gingen voorbij, maar er doken geen fossielen meer op van vroege tetrapoden. In 1982 was het al meer dan 50 jaar na de eerste ontdekking van Ichthyostega en men was geen stap verder gekomen. Over Ichthyostega was lange tijd trouwens ook weinig bekend, omdat de onderzoeker Erik Jarvik ontzettend treuzelde met het publiceren van het onderzoeksverslag. Hij publiceerde er maar sporadisch wat kleine artikeltjes over, legde het werk regelmatig neer om andere fossielen te gaan onderzoeken, waaronder van Eusthenopteron, en liet de rest van de paleontologen met hun vragen zitten. Het definitieve verslag van Ichthyostega kwam pas uit in 1996, ruim 75 jaar na de ontdekking, en twee jaar voor de dood van Erik Jarvik. In de vroege jaren tachtig hadden enkele paleontologen genoeg van het getreuzel, en ze besloten zelf eens op pad te gaan op zoek naar fossielen van vroege tetrapoden. Degene die zich dit voornamen waren de Engelse Jenny en Robert Clack, een echtpaar die beide paleontoloog waren. Gewoonlijk zochten ze fossielen in Schotse Devoonlagen, hier werden wel resten van spiervinnigen gevonden maar geen tetrapoden. Groenland leek een veel betere locatie, ten slotte kwam hier de enige gevonden tetrapode uit het Devoon vandaan. Het gebied bij Gauss HalvØ dat Sve-Sderberg en Jarvik hadden onderzocht was na hen nog n keer bezocht door een zekere John Nicholson, die in dienst was van een aardoliemaatschappij maar ook wat fossielen had beschreven en meegenomen. Volgens hem was er iets meer naar het Noorden een berg, de Stensi Bjerg, waar Devonische tetrapoden in overvloed in de rotsen staken. De Clacks kregen het met hulp van Svend Bendix-Almgreen, een Deense paleontoloog, in 1987 voor elkaar dat ze meekonden met een Deense geologische expeditie die in Groenland olie ging zoeken in het gebied waar Sve-Sderberg en Jarvik hun ontdekkingen hadden gedaan. Ze vlogen naar Groenland, en gingen per helikopter naar de kale hellingen van de Stensi Bjerg. Ze zetten hun tenten op op een hoge richel, 80 meter boven zee. Van hieruit begonnen ze de berg te beklimmen op zoek naar de plaats waar Nicholson het over had. Het beklimmen van de berg was nog niet makkelijk was voor de beide Clacks, die nog nooit eerder hadden geklommen. Onderweg speurden ze de rotsen af op zoek naar fossielen. De eerste die iets zag was Jenny Clack, die tussen de rotsen een schedel ontdekte. Bij nader onderzoek bleek het een schedel van een dier te zijn waarvan Sve-Sderberg en Jarvik tijdens de eerste expeditie enkele fragmenten hadden ontdekt. Jarvik had het dier Acanthostega genoemd, maar verder niet veel aandacht aan besteed. De schedel was duidelijk van de berghelling gerold nadat het omringende steen was weggerodeerd en verder op de helling was een spoor te zien van meer Acanthostegabotten. Ze volgden het spoor totaan de plek waar de fragmenten vandaan kwamen. Zoals Nicholson had beschreven puilden de rotsen uit van fossielen, en op de grond lagen overal brokken steen waar allerlei schedels en botfragmenten uitstaken. Ze vulden dadelijk hun rugzakken en bleven de komende twee weken terug komen tot de hele omgeving kaalgeplukt was.
De schedel van Acanthostega.
Bij elkaar hadden ze een halve ton fossielbevattende stenen verzameld, die in kisten werden verpakt en met helikopter, schip en vrachtwagen terug naar Engeland werden vervoerd. Jenny Clack richtte samen met paleontoloog Michael Coates een laboratorium op in Cambrigde, om de gevonden fossielen vrij te prepareren van de brokstukken waar ze inzaten en ze te bestuderen. Wat ze onder andere hadden gevonden waren verschillende spiervinnigen, een achterpoot van Ichthyostega, en vooral minstens 5 Acanthostegaschedels en vele andere botten. Coates was degene die begon met het te verwijderen van de steenresten van de beenderen met behulp van een pneumatisch hamertje. Hij begon met de meest veelbelovende schedel, die hij 'Boris' had genoemd. Waar hij en de Clacks op hoopte waren de vingers. De overgang van een gelobde vin naar vingers was de voornaamste verandering van vissen naar amfibien. Vijf vingers was duidelijk het standaard aantal, het was het maximale aantal vingers van tetrapoden. Sommige dieren hadden minder vingers, of zelfs helemaal geen meer, maar in zulke gevallen was door overblijvende handbeentjes duidelijk te zien welke vingers hier van het oorspronkelijke basisontwerp verdwenen waren en welke waren overgebleven. De onderzoekers hoopte vurig dat er onder het gesteente waar de Acanthostegaresten in zaten het basisgetal van vijf vingers vandaan zou komen. Coats begon met hier en daar wat sporadisch steen te verwijderen, tot hij de humerus vond, een stevig stuk bot met plaatsen voor spieraanhechtingen. Het was een humerus die heel gewoon was voor vroege tetrapoden. Hieronder zouden de radius en ulna moeten liggen. Hij maakte de humerus dus geheel schoon en begon daarna met de twee onderarmbeenderen. Met het blootleggen van de radius zag hij dat deze erg kort en dik was, en verwachte dat de ulna net zo zou zijn. Maar in plaats darvan bleek de ulna wel twee keer zo lang als de radius. Dit was op z'n minst een spectaculaire ontdekking, gewoonlijk heeft een gewerveld landdier een radius en ulna die evenlang zijn, en bij Ichthyostega is het niet anders. De overeenkomst in lengte is noodzakelijk wil het dier fatsoenlijk kunnen lopen. Maar Acanthostega had zo'n vreemd gebogen poot dat als hij er op zou gaan staan hij meteen voor -of achterover zou vallen. De vreemde verhouding tussen radius en ulna bij Acanthostega was echter identiek aan die bij spiervinnige vissen zoals Eusthenopteron.
Een deel van 'Boris' de Acanthostega. In het midden zijn de radius en ulna te zien, rechts daarvan de vingers.
Hoe langer Coats naar de poten keek, hoe minder ze geschikt bleken om op te lopen. Het schouderblad was zo dun als een eierschaal, en ook de pols was bijzonder dun, maar de radius en de ulna in de onderarm waren weer erg dik. Alles wees er op dat Acanthostega's poten niet geschikt waren om over land te lopen, maar als paddels fungeerden. Maar zoals later zou blijken was dit nog niet eens de grootste ontdekking, toen Coats verder ging met het blootleggen van de voorpoot bereikte hij de vingers. Nu Acanthostega zo'n duidelijke tussenvorm tussen vissen en amfibien representeerde verwachte hij niets anders dan onder de pols vijf vingers aan te treffen. Terwijl de avond viel ging hij door met stukjes steen wegbikken, en n voor n kwamen er weer kleine vingerkootjes tevoorschijn. Per uur verscheen er weer een nieuwe vinger, en tegen het ochtendgloren ware alle vingers blootgelegd. Maar tegen ieders verwachting in waren het geen vijf vingers, onder het steen waren er zelfs acht vandaan gekomen. Ondertussen onderzocht Jenny Clack de schedel van het dier, en ook hier kwamen weer verrassende resultaten naar voren. Om te beginnen vertoonde de schedel sporen van kieuwen, in de hals vond ze een klein staafje dat werd herkend als een kieuwboog, nou hebben moderne amfibien ook kieuwbogen, maar die dienen een ander doel, ze ondersteunen de tongspieren. Maar waar bij moderne amfibien de doorsnee van een kiewboog rond is, zijn die van vissen meer sikkelvormig, om ruimte te bieden aan bloedvaten. Acanthostega beschikt duidelijk over dezelfde kieuwboog als vissen, wat betekend dat dit dier als enige bekende tetrapode zijn hele leven lang beschikte over functionele kieuwen. Maar dat was nog niet alles, daarnaast werd ontdekt dat Acanthostega niet beschikte over het oor dat tetrapoden gewoonlijk gebruiken om boven water te horen. Vissen hebben in het geheel geen oren, maar ze kunnen geluid waarnemen door het zijlijnsysteem, waarbij de geluidsgolfjes door onderhuidse kanalen worden vervoerd. Acanthostega beschikte hier waarschijnlijk ook over, maar boven water zou hij zo goed als doof zijn geweest. Zijn gehoorsysteem was dus net als zijn ademhalingssysteem en poten aangepast aan het waterleven. Maar anders dan vissen opent Acanthostega wel een deur naar de toekomst. Aan de zijkanten van de schedel werden kleine knobbelige botstukjes ontdekt, vanuit vissenperspectief zou dit het een hyomandibulare kunnen zijn, een botstuk dat de onderkaak ondersteund en tevens helpt de kieuwkleppen te openen. Vanuit tetrapodenperspectief zou het een stijgbeugel kunnen zijn, het oorbotje dat tegen het trommelvlies drukt. Hierdoor werd bevestigd wat 19e eeuwse embryologen al hadden geconstateerd, namelijk dat het visachtige hyomandibulare homoloog is aan de stijgbeugel van tetrapoden. Hierdoor staat vast dat het kleine oorbotje zich ontwikkeld heeft uit een hulpstuk voor het bewegen van de kieuwen en het verbinden van de kaken met de achterschedel. Voor Acanthostega, die boven water doof was, had het echter nog niet de functie die het nu heeft. Pas later zou de stijgbeugel los komen van de schedel om zo geluiden door te geven aan de hersenen.
Een skelet en reconstructie van Acanthostega.
Ten slotte waren de ribben niet meer dan korte stompjes, en absoluut ongeschikt om de organen tegen de zwaartekracht te beschermen. En waar Ichthyostega alleen aan de bovenkant van zijn staart de ingewikkelde structuur van spiervinnige vissenstaart had, was de staart van Acanthostega zowel van boven als onderen identiek aan die van een spiervinnige vis. Elk lichaamsdeel wees er op dat Acanthostega een aquatisch dier was, die zich op het droge zeer ellendig zou voelen, als hij het met zijn zwakke poten al voor elkaar kreeg om uit het water te komen. Sommige lichaamsdelen waren zelfs ronduit visachtig, de structuur van de staart en kieuwen komen enkel voor bij vissen, en hoewel er verschillende amfibien zijn geweest die weer terug naar het water keerde en de kieuwen na hun larvale stadium behielden zijn dit in alle gevallen externe kieuwen, terwijl Acanthostega's kieuwen intern, zoals bij een vis zijn. En toch onderscheid dit dier zich evenwel van de vissen, misschien nog niet eens door zijn vier poten, want ook Eusthenopteron beschikt hierover, maar meer nog door de tenen. Hoewel ook deze door hun ongewone aantal weer afwijkend zijn van gewoonlijke tetrapoden. Met de publicatie van Acanthostega werden de gebruikelijke opvattingen over vroege tetrapoden totaal overhoop gegooid. Tot dan toe had men gedacht dat spiervinnge vissen eerst het land op kropen en daarna poten uit hun vinnen ontwikkelde, maar uit deze vondst bleek dat poten eerst waren ontstaan, en deze pas daarna werden gebruikt om het land op te kruipen. Hiermee werd ook aangetoond dat het ontstaan van de vier voeten en de verovering van het land geen samenhangende gebeurtenissen waren. Hoewel de ontwikkeling van de voeten een evolutionaire deur richting het land openden hadden voeten voor het zelfde geld kunnen zijn gevolueerd zonder dat er landdieren ontstonden. De vraag reist hoedanook waarom poten en vooral de tenen zijn ontstaan. Blijkbaar had Acanthostega als waterdier meer profijt van korte, gesegmenteerde vinstralen, die we nu vingers noemen, dan met de oorspronkelijke vinstralen van zijn waarschijnlijke voorouder Eusthenopteron of een verwante spiervinnige. Jenny Clacks eigen theorie is dat Acanthostega's vingers een hulpmiddel waren bij zijn manier van jagen. Net als de spiervinnige vissen en verwante amfibien was het een jager die aanviel vanuit een hinderlaag, die zich verschool tussen de waterplanten die vlak onder de oppervlakte groeide. Waarneer er een prooi voorbij kwam schoot Acanthostega met behulp van zijn krachtige zwemstaart pijlsnel naar voren, en verzwolg de prooi met zijn kaken. Maar in een hinderlaag liggen is in het water nog niet zo makkelijk. Zeker in een rivier is het water in constant in beweging, en als je niet oplet drijf je zo weg je schuilplaats uit. Wat veel moderne vissen dan ook doen is zichzelf vasthouden aan de waterplanten om te voorkomen dat ze afdrijven terwijl ze in een hinderlaag liggen. Het is maar een theorie, maar met de korte gesegmenteerde vinstralen zou Acanthostega prima in staat zijn zich vast te houden aan de vegetatie terwijl hij wachtte op een voorbij schietende vis.
Spoedig na de ontdekking van Acanthostega werd de tetrapoden collectie uitgebreid met meerdere vondsten zoals van Elginerpeton, een fossiel uit Schotland dat bestaat uit een onderkaak, een schouder, een humerus en een femur. Ook werd in Rusland een dier ontdekt dat Tulerpeton werd genoemd, en in Letland een nieuwe potentile visachtige voorouder, Panderichthys. We kunnen nu een voorlopge beschrijving geven van hoe het ontstaan van de tetrapoden en de kolonisatie van het land zich heeft voltrokken. Tenminste 337 miljoen jaar geleden ontstond er een tak van spiervinnigen die leidde naar een groep vissen met zeer stevige benige vinnen. Enkele leden hiervan waren bijvoorbeeld Eusternopteron en ook Panderichthys, deze was 60 centimeter lang, had een kop zo plat als een bijzettafeltje, en had zulke stevige vinnen dat je die rustig poten kan noemen, maar alleen wel met vinstralen in plaats van vingers. Ongeveer 10 of 15 miljoen jaar later hadden verwanten van Panderichthys hun lichaam omgebouwd tot dat van een tetrapode. Elginerpeton, een dier dat werd opgedoken uit een oude museumla is de oudste en meest primitieve viervoetige. Helaas zijn de poten voorbij de humerus en femur niet gevonden, dus is het niet na te gaan of hij vingers of vinstralen had, maar alle informatie die kan worden afgeleid uit de schaarse resten wijzen er op dat hij een tussenvorm was van spiervinnige vissen als Panderichthys en latere tetrapoden. De achterpoten waren klein en verdraaid, zodat de knien naar onderen wezen, waardoor de poten ongeschikt zouden zijn voor lopen, maar wel geschikt voor roeien. Gegeven het feit dat Elginerpeton anderhalve meter lang was en jaagde langs de bodem van rivieren was het in feite een viervoeter die leefde als een gewone vis. Binnen enkele miljoenen jaren was Elginerpeton weer verdwenen, maar overal ter wereld ontwikkelden zich nieuwe soorten tetrapoden. De familie omvatte een soort die Ventastega is genoemd, en waarvan in Letland wat schedeldelen zijn gevonden, en een reeks dieren waarvan alleen onderkaken zijn gevonden, zoals Metaxygnathus, Obruchevichthys en Densignathus. Deze vondsten zijn gedaan in verschillende delen van de wereld, en hoewel tijdens het Devoon de vorm en positie van de continenten er heel anders uit zag dan tegenwoordig lagen de plaatsen evengoed een behoorlijk eind uit elkaar. Maar dit was waarschijnlijk een afsplitsing in de stamboom die spoedig weer uitstierf, de Groenlandse Ichthyostega en Acanthostega zijn veel nauwer verwant met de latere tetrapoden. Acanthostega had vingers en tenen, een lange starre snuit die niet werd verbonden door het hyomandibulare botje. Temidden van deze reeks met visachtige tetrapoden lijkt Ichthyostega, ooit beschouwd als de directe voorouder van alle tetrapoden, niet meer dan het zoveelste natuurlijke experiment om vissen met poten uit te rusten. In sommige opzichten had hij al meer weg van de latere tetrapoden, de vissenstaart was al voor de helft verdwenen, de beenderen van zijn voorpoten waren even lang en geschikt om zijn gewicht te dragen. Ook had hij grotere ribben en de heupen hadden een kom waar de kop van de femur in paste. Toch staken de achterpoten volgens Coats en de Zweedse Hans Bjerring naar achteren zoals bij een zeehond. Ichthyostega maakte dus waarschijnlijk alleen van zijn voorpoten gebruik om het droge op te komen, terwijl de achterpoten nog alleen als paddels dienst deden in het water. In feite waren Ichthyostega en Acanthostega al in hun tijd levende fossielen, want hoewel Hynerpeton enkele miljoenen jaren ouder is staat hij wel dichter bij de latere tetrapoden. Hoewel een schouder en wat kaakdelen het enige is wat er van is overgebleven vertoond de schouder sporen van krachtigere spieren dan bij Ichthyostega. Hoe de achterpoten er hebben uitgezien is echter onbekend, en mogelijk sleepte hij deze achter zich aan zoals Ichthyostega. Tenslotte was er Tulerpeton, een dier wiens fossielen veel completer zijn dan bij de meeste andere vroege tetrapoden, zij het niet zo kompleet als Acanthostega en Ichthyostega. Grote delen van de rug en staart ontbreken, maar zowel de voor -als achterpoot zijn kompleet. Aan de voorpoot zaten 6 vingers, aan de achterpoot 7, wat suggereert dat latere tetrapoden met het standaardgetal van vijf vingers pas ontstonden met het aflopen van het Devoon. Hoogstwaarschijnlijk leefde al deze soorten met een tal van vroege tetrapoden die nog niet ontdekt zijn, maar rond 335 miljoen jaar geleden splitste twee groepen zich van elkaar af, waarbij de ene groep gestalte gaf aan de moderne amfibien, en de andere aan de amnioten, de reptielen, zoogdieren en vogels.
In het kort nog een keer; de chronologische geschiedenis:
Eusthenopteron
Tijd: ? miljoen jaar geleden Een spiervinnige vis, met in zijn vinnen typisch tetrapodische botelementen zoals de humerus, radius en ulna, je zou kunnen zeggen poten, maar dan met vinstralen in plaats van vingers. Eusthenopteron wordt beschouwd als de best gedocumenteerde gewervelde uit de Paleozosche era.
Panderichthys
Tijd: 378 miljoen jaar geleden. (Laat Givetian/vroeg Frasnian) Een spiervinnige met dezelfde botelementen als Eusthenopteron. Deze vis beschikte bovendien over een platte schedel met hoog geplaatste ogen, die zeer sterk overeen komt met die van vroege tetrapoden. Verder had hij geen rugvin, alleen 2 borst -en 2 buikvinnen, en de staartvin.
Elginerpeton
Tijd: 368 miljoen jaar geleden (Laat Frasnian) De oudst bekende tetrapode, maar door karakteristieken in de kaak is het waarschijnlijk dat Elginerpeton tot een kort levende zijtak in de tetrapoden evolutie behoord.
Hynerpeton
Tijd: 365-363 miljoen jaar geleden (Laat Famennian) Mogelijk een tetrapode waarvan in ieder geval de voorpoten aangepast waren voor leven op het land, maar dit blijft twijfelachtig door de schaarse fossielen.
Acanthostega
Tijd: 363 miljoen jaar geleden Een tetrapode die ondanks zijn goed ontwikkelde poten duidelijk niet was aangepast voor leven op het land. Als enige bekende tetrapode bezat hij interne kieuwen. Verder een staart die identiek was aan die van spiervinnigen, korte ribben, oren en een neus die geschikt waren voor onder water gebruik en 8 tenen per poot.
Ichthyostega
Tijd: 363 miljoen jaar geleden Deze was iets beter aangepast voor een leven op het land, maar kon alleen zijn voorpoten gebruiken om te lopen, de achterste sleepten er maar achteraan. Ichthyostega bezat geen interne kieuwen, en ademde wellicht door longen. Kleine karakteristieken in het skelet doen herinneren aan spiervinnige vissen.
Tulerpeton
Tijd: 363 miljoen jaar geleden Een stevige schouder suggereert dat het dier redelijk goed kon lopen. De voorpoot bevat 6 tenen, de achterpoot 7.
br />
De vinnen en voeten van (van boven naar beneden) Panderichthys, Eusthenopteron, Ichthyostega en Acanthostega. (Hu = Humerus, Ra = radius, Ul = Ulna.)
Extra toevoeging: De evolutie van longen.
Darwin, die leefde in een tijd waarin het fossielenbestand verre van kompleet genoeg was om goede uitspraken te doen over de evolutie, kon over veel diersoorten alleen wat speculeren en wetenschappelijk verantwoord gissen over de ontwikkelingen die dier -en plantensoorten hadden doorlopen. Sommige van die voorspellingen waren een schot in de roos, zo bleek later toen dankzij gevonden fossielen het plaatje stukken duidelijker begon te worden. Zo had Darwin gelijk toen hij beweerde dat de vleugels van insecten in feite hun voormalige kieuwen waren, dit is bevestigd door een aantal zeer oude fossiele insecten. Ook de voorspelling dat schakels in de menselijke evolutie voornamelijk in Afrika zouden moeten worden gevonden kwam uit. Maar uiteraard waren er ook een aantal gevallen waar hij het mis had, zijn bewering over het ontstaan van longen was een logische vergissing. Volgens Darwin, en vele wetenschappers na hem, waren longen ontstaan uit de zwemblazen van vissen. Dat leek heel logisch, de longvis van Owen liet duidelijk zien dat de zwemblaas en de longen homoloog waren, en daar vrijwel alle vissoorten over een zwemblaas beschikte moest deze worden beschouwd als een gedeeld primitief kenmerk. Vissen zouden dus de met gas gevulde zwemblaas hebben ontwikkeld om hun drijfvermogen in de hand te houden, en later, bij de spiervinnigen, zou deze zich weer ontwikkeld hebben tot longen. Meer dan een eeuw lang wist men niet beter dan dat het zo gegaan was.
Behalve steuren en ware beenvissen (90% van de huidige vissen) beschikken alle families vissen over longen in plaats van zwemblazen. (Coelacanthen hebben geen echte longen meer, maar wel rudimenten van wat longen zijn geweest.)
Maar een modern cladogram suggereert iets anders, men had gedacht dat een zwemblaas een gedeeld primitief kenmerk was omdat ongeveer 90% van alle huidige vissen er over beschikt. Kijkt men echter naar het complete aantal kenmerken dan blijkt dat de vissen met zwemblaas juist geavanceerder zijn dan die met longen en/of zonder zwemblaas. Het cladogram laat zelfs doorschemeren dat de zwemblaas is ontstaan uit longen en niet andersom! Longen zijn ontstaan als uitstulpingen van het spijsverteringskanaal dat vol met bloedvaatjes kwam te zitten. Van daaruit werden ze weer gereduceerd tot een zwemblaas, een simpele holte met gas gevuld. Maar hoe valt dat te verklaren, zijn alle zwemblaasbezittende vissen dan uit landdieren voortgekomen? Is onwaarschijnlijk... Een mogelijke verklaring ligt in het feit dat vissen zonder longen, bijvoorbeeld een forel, hun bloed rondpompen in een enkelvoudige kring. Het hart pompt het bloed richting de kieuwen, daar neemt het zuurstof op en gaat vervolgens het hele lijf af om alle spieren van zuurstof te voorzien. Echter, het hart heeft ook zuurstof nodig, en waarneer het bloed het hele lijf is langsgeweest is er nog maar weinig zuurstof over. Waarneer de vis hard moet zwemmen is dat een probleem, de staartspieren hebben meer zuurstof nodig en hoe harder de spieractiviteit, hoe minder zuurstof er over blijft voor het hart dat juist harder moet werken. Vissen zonder longen leggen na een paar minuten hard zwemmen het loodje. Een vis met longen daarentegen, bijvoorbeeld een moddersnoek, heeft het beter voor elkaar. Het hart pompt bloed naar de longen, en als het daar zuurstof heeft opgenomen gaat het direct terug naar het hart en dan pas naar de rest van het lichaam. Vissen met deze dubbele bloedsomloop hebben dus meer uithoudingsvermogen en daarmee meer evolutionair voordeel dan de longloze vissen met een enkele bloedsomloop. Er is alleen n voorwaarde aan deze constructie verbonden, namelijk dat de vis naar de oppervlakte moet om adem te halen, want longen functioneren niet onder water. Wat moderne longbezittende vissen dan ook doen is op rustige momenten adem halen met behulp van hun kieuwen, en bij drukke activiteiten zo nu en dan aan de oppervlakte adem halen zodat het hart niet overspannen raakt. Om deze reden zijn longbezittende vissen miljoenen jaren lang uiterst succesvol geweest, maar toch is er iets gebeurd waardoor longbezittende vissen nu een zeldzaamheid zijn en het overgrote deel van de moderne vissen met hun weinig geschikte zwemblazen zitten opgescheept. De verklaring hiervoor is waarschijnlijk dat vissen die zoals moderne walvissen en zeeschildpadden aan de oppervlakte lucht kwamen ademen dit veilig konden doen tot ongeveer 220 miljoen jaar geleden. Daarna werd het gevaarlijk om je als vis bij de oppervlakte op te houden, want de vliegende reptielen die we kennen als Pterosaurirs, en later zeevogels, meeuwen en dergelijke, scheerden vlak boven water en grepen elke vis die ze zagen. Wellicht is dit de reden waarom longbezittende vissen tegenwoordig bijna zijn uitgeroeid, terwijl degene die hun longen hebben omgezet in zwemblazen floreren. Zouden er nu om wat voor reden dan ook geen visetende vogels meer zijn, en ook geen andere gevaren voor een vis aan de oppervlakte, dan zouden we misschien in enkele miljoenen jaren een comeback van de longvissen, moddersnoeken en dergelijke kunnen waarnemen, doordat deze longbezittende vissen met hun dubbele bloedsomloop meer voordeel hebben dat longloze vissen.
Nog een extra toevoeging: Even de zeebenen strekken
Lange tijd was men wat betreft het ontstaan van poten uit vinnen afgegaan op de hypothese van een zekere paleontoloog Romer. Deze Romer beweerde dat vissen poten konden ontwikkelen waarneer deze bij een plotselinge droogte kwamen te stranden en dan met hun gespierde vinnen konden proberen terug te kruipen naar het dichtsbijzijnde poeltje waar nog wel water in stond. Tot zijn spijt kon Romer echter niet beschikken over fossielen om informatie uit te winnen, want de fossielen van vroege tetrapoden moesten destijds nog ontdekt worden. De later gevonden Ichthyostega leek echter wel tamelijk goed te passen binnen dit scenario. Met de vondst van Acanthostega viel de hypothese echter in duigen, want dit dier toonde aan dat poten er eerder waren dan landdieren, en bovendien heeft men aangetoond dat in het Devoon de wereld nat en moerassig was, waar geen sprake was van uitdrogingen. De conclusie was dus dat de voorouders van tetrapoden uit hun vinnen poten hadden gevormd vr ze er nog maar aan dachten om een kijkje te nemen boven water. Dan rijst er echter de vraag wat voor voordeel een waterdier kan hebben bij poten? Ik heb al eerder de hypothese van Jenny Clack vermeld, dat Acanthostega zijn vingers had om zich vast te houden aan waterplanten en zodoende er voordeel aan had, maar ik wil nog vermelden dat er meerdere voordelen zijn van poten in het water en dat deze tegenwoordig ook daadwerkelijk worden gebruikt: Het beste voorbeeld daarvan zijn de voelsprietvissen (Frogfishes in de Engelse taal):
Dit is een hele familie van afzichtelijke vissen, maar met hele bijzondere vinnen. Zoals je kan zien vertonen de orstvinnen wat overeenkomsten met poten. Er is een knie te zien, en de uiteinden van de vinstralen lijken op een soort tenen. En deze pootachtige vinnen worden ook daadwerkelijk gebruikt om over de zeebodem te lopen. Erg snel gaat het niet, amper een kilometer per uur, maar de vis heeft het voordeel geluidloos te zijn, terwijl het gebruiken van de zwemstaart gehoord zou kunnen worden door zijn prooien. Laat ik duidelijk stellen dat deze vissen geen voorouders of zelfs maar nauwe verwanten van landdieren zijn. Ze behoren tot de straalvinnigen, terwijl tetrapoden afstammen van spiervinnigen. Maar ze geven wel aan wat de redenen zouden kunnen zijn geweest voor dieren zoals Acanthostega om in het water poten uit te vinden, waaruit later bleek dat deze de dieren in staat stelden om op het droge een totaal nieuw leefgebied te exploiteren en aldus landdieren te worden.
Bronnen:
1 Aan de waterkant, Carl Zimmer 1999 De daaruit voortgekomen feiten worden bevestigd door primaire literatuur van Clack, Coats, Jarvik, enz.
Is ID de wetenschappelijke (sic) uitleg voor creationiisten 150jaar Darwinisme... De evolutietheorie gewikt en gewogen David sorensen en revolutietheorie lalala DE VERRIJZENIS VAN DE JONGE VERDRONKEN KOE » Reageer (21) ID TOVERDOOS OVER SLECHTE ID-EETJES ,RESISTENTIE & TOVERDOCTORS c) OEC d) YEC
Een oeroud spoor Denisova : ZUID SIBERIË DENISOVA - mens
DMANISI AAP OF MENS DE OUDSTE ? » Reageer (1)Nakalipithecus nakayamai (<) Alweer eentje ? ..... De tand des tijds / Heidelberg-mens DE HEUPEN VAN EVA CASABLANCA MAN /erectus Hobbit is aparte soort ? » Reageer (2) NOG EEN BENDE BIJTERS POLONAISE met neanderthaler TANDEN UIT DE QESEM GROT Neanderthaler genoom Mitochondriale Genenkaart van Neanderthaler Xuchang mens
b)Biologie 1.-(EVOLUTIE ) NAS / IM Document 2008 (2) NAS / IM Document 2008 ( 1)
Cambrium & precambrium AVALON GABONESE chips
Evolutie in actie Opmerkelijk snelle adaptaties bij kroatische ruine-hagedissen:
EEN PLUIM VOOR CHINA Geef eens een pootje Pluimgewicht KLEIN DUIMPJE & DE REUS &VEREN Opnieuw gevederde Maniraptor Similicaudipteryx. Aerosteon riocoloradensis
Krokodillen
PAKASUCHUS KAPILIMAI Prestosuchus chiniquensis Terug naar zee viseter
Eritherium azzouzorum LJOEBA Darwinius masillae VLIEGENDE KATTEN ? Vleermuizen ALWEER EEN BELANGRIJK STUK UIT DE LEGPUZZEL Zee-zoogdieren ; Van de wal in de visgronden Indohyus / walvisevolutie Zeehonden-evolutie
*Blauwe en groene ( =nederlandse)teksten zijn meestal aanklikbare links *Engelse Wikipedia teksten verwijzen in de linkerkolom naar verschillende niet-engelse versies van het wikiartikel *Blauwe teksten tusssen "" , zijn voornamelijk ( gedeeltelijke)citaten afkomstig van mensen met andere meningen
Evolutie / Charles Darwin aan de basis: SELECTIE NATUURLIJK DE LEVENSBOOM UITSTERVEN DEEP TIME Biogeografie SEKSUELE SELECTIE Al jaren lang samen CO-EVOLUTIE De economie van de natuur Geleidelijke veranderingen
Unintelligent design:
IP Theorie Ons gebit , De appendix, Geboorte, De weg van het zaad , Nervus Laryngeus Recurrens, DNA , 75 mistakes , Het boek
Unintelligent design (1): Rechtop lopen Tomaso over het oog , oogspieren , Unintelligent Design (5): ZWEETVOETEN Unintelligent design (6): Vitamine C Unintelligent design (8): Kuitspieren
 Unintelligent design (1): Rechtop lopen
Unintelligent design (1): Rechtop lopen

)

 artistieke reconstructie
artistieke reconstructie ")














 (artist reconstruction Ted Daeschler)
(artist reconstruction Ted Daeschler)






)










 fossiele schouder van
fossiele schouder van  fossiele kaak van
fossiele kaak van 
 fossielekaak van
fossielekaak van 




































 Deze metapterygiale as is duidelijk terug te vinden in de vin van Eusthenopteron, en ook hier zijn de vinstralen vertakkingen van de as, net zoals de vingers van een tetrapode embryo. In dit licht kun je constateren dat de vinnen van Eusthenopteron ineengedrukte handen en voeten zijn. Of beter gezegd, onze handen en voeten uiteengetrokken vinnen zijn met gesegmenteerde vinstralen.
Deze metapterygiale as is duidelijk terug te vinden in de vin van Eusthenopteron, en ook hier zijn de vinstralen vertakkingen van de as, net zoals de vingers van een tetrapode embryo. In dit licht kun je constateren dat de vinnen van Eusthenopteron ineengedrukte handen en voeten zijn. Of beter gezegd, onze handen en voeten uiteengetrokken vinnen zijn met gesegmenteerde vinstralen.


















 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}