Dit blog is een onderdeel van EVODISKU WAT IS DE BEDOELING EN WAAR STAAT DIT BLOG VOOR
****Wie meent dat alles inmiddels wel over de evolutietheorie gezegd is en dat de discussie gesloten kan worden, ziet over het hoofd dat de wetenschap niet stil staat.
***Wie meent dat inmiddels het creationisme definitief het pleit heeft verloren en dat de discussie gesloten kan worden , ziet over het hoofd dat het "creationisme" is geevolueerd ( en zal evolueren ) in nieuwere mimicrytische vormen( meme-complexen ) zoals bijvoorbeeld het ID(C)
***Dit blog is speciaal opgezet om de aktualiteit binnen het evolutie-creationisme debat te volgen en van kommentaren te voorzien ... waartoe de lezers zijn uitgenodigd bij te dragen ...
Let echter wél op het volgende :
"Je bent een rund als je hier met religie stunt " ....
09-10-2009
Wie oren heeft , die hore
Zoogdieren en middenoor Er is weer een zoogdier gevonden uit het reptielen tijdperk Het fossiel is afkomstig van de beroemde vindplaatsen in de Yixian formatiesin het NO van China ...
Chinese en Amerikaanse paleontologen publiceerden in het wetenschappelijke tijdschrift Science een artikel over een fossiele insecteneter. De ouderdom van het zoogdier wordt geschat op 123 miljoen jaar, midden in het Mesozoïcum, het tijdperk van de dinosauriers
Wat ons het meest interesseerde , is het oor van deze insecteneter, zei onderzoeker dr. ZheXi Luo, van het Carnegie Museum of Natural History. Zoogdieren hebben een veel gevoeliger gehoor dan andere gewervelde dieren.
MIDDENOOR
Zoogdieren hebben een veel beter ontwikkeld gehoor dan alle andere gewervelde dieren, en mede daardoor konden de zoogdieren een leefwijze ontwikkelen die sterk op dat goede gehoor berust.
Evolutiebiologen veronderstellen dat de ontwikkeling van een complexe en efficient gehoor een overlevingsvoordeel bood ; detectie van prooien en praedatoren werd vergemakkelijkt en de communicatie ( dus ook het vinden van een geschikte partner ) verbeterd
Dankzij de gevoelige middenoorstructuur hoort een zoogdier zowel hoge als lage tonen. Het zoogdier-middenoor, bestaat uit drie botjes (stijgbeugel, hamer en aambeeld) en een benige ring voor de trommelholte.
Juist daarom hebben paleontologen en evolutionaire biologen al langer dan een eeuw naarstig gespeurd naar aanwijzingen die zouden kunnen leiden tot inzicht in de evolutionaire ontwikkeling van het oor.
aantoonbare/waargenomen anatomisch tusenstappen in de middenoorstructuur van fossiele "primitieve " zoogdieren in elkaar zaten , bleef tot en met de ontdekking van Yanacodon onduidelijk....
De in 2007 ontdekte Yanocodon liet voor het eerst zo'n tussenliggende stap zien. Daar is nu de maotherium asiaticus bij gekomen
Vergelijking tussen het oor en de kaak van Yanocodon & Maotherium met die van verwanten
All modern mammals (platypus, opossum and human) have a middle ear separated from the lower jaw (see example from living opossum). This jaw-ear separation is an important evolutionary innovation. It becomes possible for mammals to have a delicate and highly sensitive ear structure for better hearing, and to have a more robust lower jaw and jaw hinge for better feeding. Also, the jaw and ear are not interfering with each other. By comparison, the middle ear bones are a part of the lower jaw and form the jaw hinge in pre-mammalian relatives (see the example of Morganucodon).
The ear bones in Maotherium are partly separated from the jaw, and more similar to those in modern mammals than to mammaliaforms, but still retain the pre-mammalian condition in which the jaw and the ear are connected to each other. Moreover, the connected jaw-ear structure of Maotherium is similar to the ear structure of modern mammals at embryonic and fetal growth stages. This phenomenon is known as paedomorphosis (paedo: child-like; morphosis: similarity) and is caused by timing change in growth. The analysis of the new fossil suggests that the evolutionary pattern of the mammalian ear is directly related to timing changes in growth, as well as in changes in genes for mammalian development.
(Photo Credit: Zhe-Xi Luo/Carnegie Museum of Natural History)
Maotherium is related to such modern mammals as Didelphis. But unlike marsupials and placentals, in which the middle ear is separated from the mandible, in Maotherium the middle ear is still attached to the mandible. Recent developmental biology studies have shown that the connection of middle ear to the mandible can re-appear as the genes control their development can change, and such genetic and developmental changes can impact evolution. Maotherium provides a strong case of how development has impacted fossil evolution in the deep history of Earth. Credit: Zhe-Xi Luo/Carnegie Museum of Natural History
De middenoorbotjes van Maotherium asiaticus zijn voor het grootste deel gelijk aan die van hedendaagse zoogdieren. Volgens de onderzoekers heeft het dier daarnaast een ongebruikelijke verbinding tussen de onderkaak en het middenoor, het verbeende Meckels kraakbeen. Deze structuur komt voor bij zoogdierembryos en in het middenoor van uitgestorven zoogdieren.
Fossiel zoogdier met unieke eigenschappen toont evolutie van middenoor Auteur: prof. dr. A.J. (Tom) van Loon
De vondst van een zoogdier in 2007 , dat 125 miljoen jaar geleden leefde in wat nu de Chinese provincie Hebei is, toonde voor het eerst hoe het middenoor - een van de meest kenmerkende verschijnselen van de moderne zoogdieren - zich evolutionair ontwikkelde.
Het dier, dat Yanocodon allini is genoemd, werd ongeveer 300 km van Beijing gevonden in de Yixian-Formatie; het is het eerste fossiele zoogdier dat in de provincie Hebei is gevonden. Het dier, dat 15 cm lang was en waarschijnlijk ongeveer 30 gram woog, had tanden die kenmerkend zijn voor dieren die insecten en wormen eten. Met zijn lange lichaam en korte poten met klauwen was hij goed toegerust om te graven of onder de grond te leven.
Het holotype van Yanocodon allini, aangetroffen op een splijtvlak waardoor het fossiel in beide stukken kan worden bestudeerd.
Bij onderzoek bleek dat het middenoor van Yanocodon het midden houdt tussen dat van de moderne zoogdieren en dat van de naaste verwanten; het biedt volgens de onderzoekers een zeldzame mogelijkheid om de ontwikkeling van het zoogdieroor te onderzoeken. Dat is niet alleen interessant omdat daarmee een van de specifieke evolutionaire aanpassingen van zoogdieren kan worden onderzocht, maar ook omdat daaruit blijkt hoe een complexe structuur geleidelijk door evolutie ontstaat (en niet door intelligent design zoals de creationisten beweren(= een IC ontwerp ).
Vindplaats van Yanocodon
Reconstructie van het skelet (door Mark A. Klingler, CMNH)
Daarnaast vertoont Yanocodon nog enkele unieke eigenschappen. Zo had hij verbazend veel wervels: 26 in zijn borst en heup, terwijl de meeste uitgestorven zoogdieren er slechts 19 of 20 hadden. Mede aan dat grote aantal wervels dankt hij zijn langgerekte vorm. Een andere zeldzame eigenschap is dat hij ook ribben ter hoogte van zijn middel had. Met al deze kenmerken lijkt Yanocodon soms dichter bij de Marsupialia (buideldieren) te staan dan bij de Placentalia (zoogdieren die bij zwangerschap een placenta ontwikkelen), soms juist andersom.
Reconstructie van Yanocodon allini (door Mark A. Klingler, CMNH).
Referenties:
Luo, Z.-X., Chen, P., Li, G. & Chen, M., 2007. A new eutriconodont mammal and evolutionary development in early mammals. Nature 446, p. 288-293.
* Hieronder heb ik, integraal een encyclopedisch artikel geplaatst van de univ. van California http://beta.revealedsingularity.net/article.php?art=mammal_ear Het artikel toont de fossiele "overgangen " aan die hebben geleid tot de uiteindelijke ontwikkeling van het huidige midden-oor-apparaat bij zoogdieren
Evolutie van het zoogdieren-middenoor
De evolutie van het zoogdieren middenoor is een uniek gebeuren binnen de tetrapoden . De drie gehoorbeentjes van het zoogdieren middenoor, zijn in feite een Synapomorfie / Synapomorphy/
Het is een veel gemaakte misvatting dat de transities , die uiteindelijk uitkwamen bij de "middenoorbeentjes van zoogdieren, "niet zou zijn voldoende gedocumenteerd en wél omdat de "middenoor"- beentjes eigenlijk veel te klein en te bros zijn om te worden bewaard en/of te worden achterhaald in het fossielen bestand ...zodat er dus geen feitelijk materieel bewijs( fossiele evidenties ) is voor deze transities
Figure 1. Cladogram with representative organisms showing the changes in relative position of the quadrate, articular, squamosal and dentary in the synapsid line. The outgroup used here is a tuatara (Sphenodon, Rhynchocephalia), which has a less heavily modified skull.
Mammalian and non-mammalian jaws. In the mammal configuration, the quadrate and articular bones are much smaller and form part of the middle ear. Note that in mammals the lower jaw consists of only the dentary bone. (wikipedia )
All non-synapsid
1.- *Lineage of tetrapods containing mammals 2.- *"Lizard-faces", including crocodiles, turtles, birds, and lizards
Amniotes have a jaw joint that has an articulation between the Quadrate bone and the articular bone( at the rear end of the lower jaw.) While many sauropsids have heavily modified their skulls[1], this two-bone articulation point is still readily observable1[2]. However, it can be seen that through a rather gradual process the point of articulation in synapsids grew to accommodate a two-joint articulation which finally became the middle-ear / jaw system in extant mammals. This, in turn, could grant them higher auditory sensitivity than is found in all other tetrapods, which only have the stapes[3] and lack the mammalian incus and malleus.
Looking at Figure 1, it can be seen that early synapsids, sauropsids, and thus the common ancestor of both employed an articulation point comprised of the quadrate and articular bones. However, even in early specimens such as Procynosuchus, the quadrate and articular are both strongly reduced
Figure 2. Varanid and snake skulls. Note the articulation between the articular and quadrate in the varanid, and the compound and quadrate in the snake. Modified with permission.
with respect to extant sauropsids. By the arrival of Probainognathus, the synapsid jaw articulation had been highly modified. Many of the various other jaw elements seen in Procynosuchus had fused into the dentary bone, and the articular and quadrate had been further reduced. For this animal, all four bones were articulating with each other, reducing the reliance on any individual bone (or pair of bones) for the articulation, allowing further modification of the jaw joint. There are other animals, such as Diarthrognathus, for which this articulation is even more evident2.
Upon the arrival of animals such as Morganucodon, the quadrate and articular were barely involved in the articulation, to the point where they become nearly invisible for diagrammatic purposes. This condition is then easily modified into the solely dentary-squamosal articulation present in mammals[4], represented in Figure 1 by a possum.
Figure 3. Diagram of the middle ear from Gray's Anatomy. Image taken from the public domain.
In modern mammals, the articular and quadrate reside in the middle ear, connected to the stapes in series and have lost their connection to the jaw, which is composed in mammals of only the dentary bone. This modification to the jaw makes it more robust, but the tradeoff is that mammals have less flexibility in their jaw movements and can never move their jaws in the way that a snake or monitor lizard might.
Simple presence of a change is insufficient, however, and the selective pressures for causing such a change are important to recognize. In dicynodonts, therocephalians, and cynodonts vibrations from the air passed primarily through the mandible,
Lower jaw. Just the dentary in mammals, but the collection of bones comprising the lower jaw in all other animals
to the quadrate, to the stapes.
The "stirrup" in humans
By loosening the bones
posterior
Toward the back of the animal
to the dentary, they became more free to vibrate and thus could transmit higher frequencies3. Once these became connected with the stapes[5], they permitted not only the hearing of higher frequencies but of lower amplitudes, as well, acting as an amplification system. The resulting excellent hearing fits with fossil finds that suggest Mesozoic mammals were primarily burrowing, climbing, and/or insect eating4. A plausible and generally the most well-accepted explanation for this series of adaptations being selectivly advantageous is that it permitted these Mesozoic premammalian ancestors to better hear insects and other small prey, most particularly during the night. This works with the fossil evidence and other hypotheses about
endothermy,
A more rigorous term for "warm-bloodedness"
molar shapes, and generally unremarkable mammalian color vision to conclude mammal ancestry is rooted in nocturnal insectivorous niches, because most other terrestrial niches were occupied by dinosaurs[6].
Philip Kahn University of California, Berkeley Submitted August 17, 2008
[1] Particularly among varanids and snakes due to cranial kinesis
[2] In snakes, the lower jaw bones have been fused into the dentary and the "compound bone" which includes the articular
[3] Known as the "stirrup" in humans
[4] Needless to say, the precise transformation of the ancestral jaw joint to the mammalian middle ear is more complicated than presented, and more detail can be found in the cited article by Luo (2007)
[5] In the ear, these bones are known as the incus and malleus, respectivley
[6] While there may be dinosaurs that were nocturnal and/or insectivorous, they were probably less of a direct threat to the Mesozoic synapsids
Pough et. al. 2004. Herpetology. ISBN 0-13-100849-8
Kitzmiller v. Dover trial testimony
Allin 1975. Evolution of the mammalian middle ear. Journal of Morphology.
Luo 2007. Transformation and diversification in early mammal evolution. Nature.
Castorocauda lutrasimilis /2006 http://www.sciencedaily.com/releases/02/060224195600.htm Onderzoek in China heeft in 2006 een bijzonder fossiel opgeleverd. Het gaat om de 164 miljoen jaar oude restanten van een primitief zoogdier uit de Jiulongshan Formatie in Binnen-Mongolië. Uit een reconstructie blijkt dat dit waarschijnlijk het oudst bekende zwemmende zoogdier moet zijn geweest.
Het dier behoort tot de docodonten, een inmiddels uitgestorven groep van zoogdieren die voor zover bekend leefden van het Midden-Jura tot het Laat-Krijt, en die niet direct verwant zijn aan de moderne zoogdieren.
De artistieke weergave (links) van Castorocauda lutrasimilis (Mark A. Klinger, CMNH) op basis van o.a. het gevonden skelet (rechts)
Holotype of Castorocauda lutrasimilis [Jinzhou Museum of Paleontology (JZMP) 04-117]. (A) Photograph of the holotype. (B) Osteological structures and preserved soft-tissue features. Abbreviations: as, astragalus; ca, caudal vertebrae; cn, ento-, meso-, and ecto-cuneiforms; co, coronoid process of dentary; cp, carpals; cs, calcaneus; ec, ectepicondyle and supinator shelf (humerus); ef, entepicondyle foramen; ep?, probable epipubis; is, ischium; J, jugal; L1-6, lumbar ribs 1 to 6; m, molars; mb, manubrium of malleus; mp, metacarpals; mx, maxilla; px, premaxilla; ra, radius; rc, radial condyle; S1-2, sacrals 1 and 2; sp, extratarsal ("poisonous") spur; t4-t14 (preserved ribs through thoracic 17); uc, ulnar condyle; ul, ulna.
Het fossiel / http://scienceblogs.com/pharyngula/2006/02/jurassic_beaver.php
Het goed bewaarde fossiel is Castorocauda lutrasimilis genoemd (çastoro is het Latijnse woord voor bever; cauda het Latijnse woord voor staart, lutra het Latijnse woord voor rivierotter, en similis is het Latijnse woord voor gelijkend). Het gaat dus om een dier dat lijkt op een rivierotter, met een beverstaart. De restanten tonen aan dat het skelet en de zachte weefsels het dier goede zwemeigenschappen gaven, en de tanden tonen aan dat het om een viseter ging. Het dier moet een dichtbehaarde pels hebben gehad, waarmee het het oudste zoogdier is waarvan vaststaat dat hij een haardos had. Hij had een dichte, korte ondervacht, die zijn huid beschermde tegen direct contact met water, met daaroverheen een langere vacht.
De levenswijze moet volgens de onderzoekers geleken hebben op die van een vogelbekdier. De meest waarschijnlijk leefomgeving was langs rivieren of meren. Daar peddelde hij rond, min of meer zoals honden dat nu doen, at waterdieren en insecten, en groef gangen om in de bodem een nest te bouwen. De beverachtige staart moet geholpen hebben bij de voortbeweging in het water, net zoals bij bevers. Daarop wijzen ook de botten in zijn staart, waarvan de bouw grote gelijkenis vertoont met die van recente bevers en otters.
Vindplaats van het fossiel.
De meeste Mesozoïsche zoogdieren waren klein (met een gewicht van minder dan 50 g), en ze waren terrestrisch (op het land levend). Vrijwel allemaal voedden ze zich met insecten. Castorocauda is met zijn lengte (zonder staart) van ten minste 42,5 cm (waarvan de schedel 6 cm uitmaakte), een geschat gewicht van 500-800 g en een dieet van waterdieren daarop een duidelijke uitzondering.
Referenties:
Martin, Th., 2006. Early mammalian evolutionary experiments. Science 311, p. 1109-1110. Ji, Q., Luo, Z.-X., Yuan, C.-X. & Tabrum, A.R., 2006. A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science 311, p. 1123-1126.
Fotos welwillend ter beschikking gesteld door Zhe-Xi Luo, Carnegie Museum of Natural History (CMNH), Pittsburg, PA (Verenigde Staten van Amerika).
Het eerste in China ontdekte "oudste "zoogdier van de groep, is een kleine, op een huismuis lijkende diersoort met de naam Eomaia scansoria (vrij vertaald: oude moeder). Het eerder goed bewaard gebleven fossiel van dit diertje werd gevonden in China. Vindplaats is de beroemde Yixian formatie waar ook overblijfselen van gevederde dinosauriërs werden gevonden.
Het bijzondere van deze vondst is de goede staat waarin het (bijna complete) fossiel verkeert. Het fossiel is zo goed bewaard gebleven dankzij het vulkanische materiaal waarmee het werd bedekt. Botten van handen en voeten en een vacht waarin ten minste twee verschillende haartypen voorkomen kunnen worden onderscheiden.
Eomaia scansoria gevonden in sedimenten van de Yixian formaties in China. (bron: Carnegie Museum of Natural History)
Eerder gevonden resten van prehistorische zoogdieren bestonden slechts uit stukjes gebit. De onderzoekers hebben daarom een goed beeld van de manier waarop het diertje geleefd moet hebben. De vorm van de poten, afmetingen van de ledematen en lange vingers en tenen wijzen op een behendige klimmer.
Het ongeveer 16 cm lange diertje heeft meer overeenkomsten met de huidige placentale zoogdieren (inwendige dragers) dan met de buideldieren. Het oudste tot nu gevonden fossiel van een placentaal zoogdier had een ouderdom van 110 miljoen jaar. Deze vondst lijkt er nu op te wijzen dat de het oudste placentale zoogdieren minstens 125 miljoen jaar oud is. De splitsing tussen buideldieren en placentale zoogdieren heeft dus voor het ontstaan van Eomaia plaatsgevonden.
Alhoewel Eomaia waarschijnlijk niet de directe voorouder van alle(huidige ) zoogdieren is kan deze soort worden gezien als een zeer verre oom of tante van de mens.
Eomaia scansoria (Chinese Academy of Geological Sciences (CAGS) 01-IG-1a, b; holotype). a, Fur halo preserved around the skeleton (01-IG-1a, many structures not represented on this slab are preserved on the counter-part 01-IG-1b, not illustrated). b, Identification of major skeletal structures of Eomaia. c, Reconstruction of Eomaia as an agile animal, capable of climbing on uneven substrates and branch walking. Taken from ref (1)
150 miljoen jaar geleden werd de kunst van het termieten-eten voor het eerst uitgevonden.
Fruitafossor windscheffelia heet het beestje dat Zhe-Xi Luo en John Wible deze week in Science beschrijven. Het diertje ter grootte van een eekhoorn struinde rond op aarde toen de dinos het hier voor het zeggen hadden, zo'n 150 miljoen jaar geleden aan het van het tijdperk Jura.
Het bijzondere aan Fruitafossor is dat zowel zijn tanden als zijn pootjes lijken op die van hedendaagse termieteneters, zoals miereneters, aardvarkens en het gordeldier. Zo zijn de tanden van Fruitafossor hol, en bevatten ze geen laagje loeihard email. Kauwen kan je er niet mee, maar termieten met kop en staart wegslikken gaat uitstekend. Zo doen aardvarken, gordeldier en miereneter het immers ook, met een listige beweging van de tong.
Met zijn voorpootjes groef het dier waarschijnlijk termietenheuvels af, net zoals de hedendaagse insekteneters. Opmerkelijk is dat die laatste geen verwanten zijn van Fruitafossor ze ontstonden pas 100 miljoen jaar later. Termieteneters zijn dus zeker twee keer in de evolutie opgedoken, een verschijnsel wat biologen convergente evolutie noemen.
Oudste placentalia ?
2007 / Wible vond een nieuw zoogdierfossiel dat een placenta had.
Vergelijking van de schedels van Maelestes gobiensis (links) = De schedel die Wible en zijn collega's gebruikten in hun berekeningen en Parascalops breweri, een nu in Amerika voorkomende mol (rechts). de papierclip is van het kleinere soort 2,5 cm
De schedel van Maelestes gobiensis (zijaanzicht) / Een schets van de schedel.
Hij besloot ermee een grote berekening te maken, waarin hij andere fossielen en levende dieren meenam. De methode van Wible gaat als volgt. Hij deelde alle uiterlijke kenmerken, zoals hoektanden, in heel erg gedetailleerde categorieën in. De uiterlijke verschillen tussen dieren van nu zijn groter dan tussen de fossielen. Wible rekende terug naar het punt waar het verschil ophield. En voilà, het eerste placentale zoogdier. Deze anonieme voorouder zou 65 miljoen jaar geleden hebben rondgestruind. Dat rijmt aardig met de oudst gevonden verwant: het fossiel van een konijnachtig dier dat 63 miljoen jaar terug met moederkoek rondhuppelde Maar of er geen oudere zijn ?
Nieuw fossiel wijst op ontstaan van zoogdieren met placenta omstreeks K/T-grens
De zoogdieren bestonden, zoals tal van fossiele vondsten uitwijzen, al in het Mesozoïcum. Binnen de groep van de zoogdieren behoren de meeste soorten (5080 van de huidige 5416 bekende soorten) tot de Placentalia (zoogdieren waarvan het vrouwtje de foetus voedt via een placenta); daarnaast bestaan er nog buideldieren (Marsupialia) en enkele eierleggende zoogdieren (Monotremata).
Over het ontstaan van de Placentalia bestaat onenigheid. Volgens paleobiochemici zou die groep al voor de K/T-grens zijn ontstaan, volgens DNA-onderzoek 129-78 miljoen jaar geleden.
Opvallend genoeg zijn er echter nooit Mesozoïsche zoogdieren gevonden waarvan duidelijk was dat het Placentalia waren (maar zulke vondsten zijn er wel uit het begin van het Tertiair), dus veel paleontologen denken dat de groep omstreeks 65 miljoen jaar geleden moet zijn ontstaan.
De schedel van Maelestes gobiensis (onderaanzicht)
De vondst in Mongolië van een goed bewaard zoogdier lijkt het pleit in het voordeel van de paleontologen te beslechten. Het opvallende is dat deze conclusie niet eens direct voortkomt uit de vondst zelf, maar een gevolg is van onderzoek dat is uitgevoerd naar aanleiding van de vondst.
De onderzoekers wilden van het fossiel, dat werd gevonden in de 75-71 miljoen jaar oude Djadokhta-Formatie (en dat dus van 10 miljoen jaar voor de K/T-grens stamt), namelijk zijn juiste plaats geven binnen de zoogdieren.
Daarom deden ze een uitgebreid onderzoek naar de specifieke kenmerken van 69 taxa van de drie hoofdgroepen van uitgestorven en nog levende zoogdieren. Daarbij ging het om niet minder dan 409 anatomische karakteristieken. Bovendien onderzochten ze alle zoogdierrestanten van enige betekenis die bekend zijn uit het Krijt.
Het nieuw gevonden zoogdier zou behoren tot een tak die was afgesplitst van de omstreeks 125 miljoen jaar geleden ontstane Eutheria. Het zou nog niet tot de Placentalia hebben behoord, want die zouden pas kort na de K/T-grens zijn ontstaan.
Volgens deze zienswijze, die op veel overtuigende gegevens is gestoeld maar die ongetwijfeld veel kritiek zal krijgen omdat eerdere stambomen behoorlijk worden veranderd, is het oudst bekende zoogdier dat tot de Placentalia gerekend mag worden een konijnachtig dier uit Azië van ongeveer 63 miljoen jaar oud.
De plaats van Maelestes binnen de zoogdieren; het gele gebied bestrijkt de Placentalia
Simplified eutherian part of the strict consensus of three most parsimonious trees (2,296 steps; see Supplementary Information). Broken line, K/T boundary at 65 million years. Filled circle taxa, Laurasian (northern continents); open circle taxa, Gondwanan (southern continents). Circles and circles with thicker black lines indicate temporal occurrence of studied specimens. The grey box delimits Placentalia, the oldest member of which is the early Palaeocene Mimotona. Placentalia originates at or near the K/T boundary in Laurasia. Deccanolestes from the latest Cretaceous of India is nested among Asian clades and is more parsimoniously interpreted as an independent southern migration. The animal silhouette spotlights Maelestes.
Dat alles naar aanleiding van de vondst van een incomplete schedel, wat wervels, een paar incomplete ribben en nog wat botten.
De schedel vertoont enige gelijkenis met die van een recente mol, Parascalops breweri, maar had meer ruimte voor zijn tanden. Het dier was klein (de schedel is ongeveer 2,5 cm lang), zoals overigens vrijwel alle vroege zoogdieren. De onderzoekers hebben het dier Maelestes gobiensis gedoopt.
J. R. Wible, G. W. Rougier, M. J. Novacek & R. J. Asher Nature 447, 1003-1006(21 June 2007) doi:10.1038/nature05854
Left panel: reconstructions of Maelestes, Asioryctes and Zalambdalestes, the last two of which are redrawn from ref. 15. Right panel: incomplete Maelestes skull in left lateral (top) and ventral (bottom) views, and left Maelestes mandible in lateral view (middle). The large opening in the palate between the palatines and maxillae is a palatal vacuity. Maelestes probably had some upper incisors, but only a small non-tooth-bearing fragment of the premaxilla is preserved. Scale bars, 5 mm.
Uppers are shown in labial (a) and occlusal (b) views; lowers are shown in occlusal (c) and labial (d) views.
The Fossil Record: Evolution or "Scientific Creation"
Clifford A. Cuffey
Mammal-Like Reptiles
As previously stated, a succession of transitional fossils exists that link reptiles (Class Reptilia) and mammals (Class Mammalia). These particular reptiles are classifie as Subclass Synapsida. Presently, this is the best example of th e transformation of one major higher taxon into another. The morphologic changes that took place are well documented by fossils, beginning with animals essentially 100% reptilian and resulting in animals essentially 100% mammalian. Therefore, I have chosen this as the example to summarize in more detail (Table 1, Fig. 1).
Skulls and jaws of synapsid reptiles and mammals; left column side view of skull; center column top view of skull; right column side view of lower jaw. Hylonomus modified from Carroll (1964, Figs. 2,6; 1968, Figs. 10-2, 10-5; note that Hylonomus is a protorothyrod, not a synapsid). Archaeothyris modified from Reisz (1972, Fig. 2). Haptodus modified from Currie (1977, Figs, 1a, 1b; 1979, Figs. 5a, 5b). Sphenacodo nmodified from Romer & Price (1940, Fig. 4f), Allin (1975, p. 3, Fig. 16);note: Dimetrodonsubstituted for top view; modified from Romer & Price, 1940, pl. 10. Biarmosuchus modified from Ivakhnenko et al. (1997, pl. 65, Figs. 1a, 1B, 2); Alin & Hopson (1992; Fig. 28.4c); Sigogneau & Tchudinov (1972, Figs. 1, 15). Eoarctops modified from Broom (1932, Fig. 35a); Boonstra (1969, Fig. 18). Pristerognathus modified from Broom (1932, Figs 17a, b,c); Boonstra (1963, Fig. 5d). Procynosuchus modified from Allin & Hopson (1992, Fig. 28.4e); Hopson (1987, Fig. 5c); Brink (1963, Fig. 10a); Kemp (1979, Fig. 1); Allin (1975, p. 3, Fig. 14). Thrinaxodon modified from Allin & Hopson (1992, Fig. 28.4f);Parrington (1946, Fig. 1); Allin (1975, p. 3, Fig. 13). Probainognathus modified from Allin & Hopson (1992, Fig. 28.4g); Romer (1970, Fig. 1); Allin (1975, p. 3, Fig. 12). Morga nucodon modified from Kermack, Mussett, & Rigney (1981, Figs. 95, 99a; 1973, Fig. 7a); Allin (1975, p. 3, Fig. 11). Asioryctes modified from Carroll (1988, Fig. 20-3b). Abbreviations: ag = angular; ar = articular; cp = coronoid process; d = dentary; f = lateral temporal fenestra; j = jugal; mm = attachment site for mammalian jaw muscles; o = eye socket; po = post orbital; q = quadrate; rl = reflected lamina; sq = squamosal; ty = tympanic.
TAXONOMY
LATERAL TEMPORAL FENESTRA
LOWER JAW DENTARY
TEETH
LOWER JAW: POST- DENTARY BONES
MIDDLE EAR & JAW ARTICULATION
M: Early Placental mammals Asioryctes Upper Cretaceous
Merged with eye socket; cheek arch bowed out laterally
100% of jaw length is the den- tary; condylar process in contact with squamosal
Fully differentiated teeth; incisors, canines, premolars; one tooth replacement
Much larger than eye socket; 40- 45% of skull length; expanded posterioirly, medially, & laterally; midline of skull narrow sagittal crest; chek arch bowed out laterally
95% of jaw length is the dentary; large coronoid process expanded posteriorly; condylar process expanded posteriorly
Large single canine; cheek teeth multicusped; tooth replacement reduced
20% of jaw length; angular notch widened ventrally; width of main part of angular decreased; reflec - ted lamina decreased to narrow ribbon-like horseshoe
Stapes extends from inner ear capsule to quadrate; quadrate tiny; quadrate-articular joint
I:Galesaurid cynodonts Thrinaxodon Lower Triassic
Much larger than eye socket; 40% of skull length; expanded pos- terioirly, medially, & laterally; midline of skull narrow sagittal crest; chek arch bowed out laterally
85% of jaw length is the dentary; large coronoid process expanded to top of eye socket and pos- teriorly; jaw muscles attached to most of coronoid process
Large single canine; cheek teeth multicusped; tooth replacement reduced
25% of jaw length; angular notch widened ventrally; width of reflec- ted lamina decreased; width of main part of angular decreased
Stapes extends from inner ear capsule to quadrate; quadrate small; quadrate-articular jaw joint
Much larger than eye socket; 40% of skull length; expanded pos- terioirly, medially, & laterally; midline of skull narrow sagittal crest; chek arch bowed out laterally
75-80% of jaw length is the den- tary; coronoid process expanded to near top of eye socket and posteriorly; jaw muscles attached to dorsal part of coronoid process
Large single canine; cheek teeth multicusped
30% of jaw length; angular notch widened ventrally; width of reflected lamina decreased
Stapes extends from inner ear capsule to quadrate; quadrate small; quadrate-articular jaw joint
G: Early Therocephalians Pristerognathus lower Upper Permian
Larger than eye socket; expanded posteriorly and medially; 30% of skull length
75-80% of jaw length is the den- tary; posterior end of dentary expanded posteriorly and dorsally into narrow blade-like coronoid process; rises to middle of eye socket
Large single canine; other teeth simple cones.
35% of jaw length; angular notch deepened into a cleft; reflected lamina large, broad, blade-like
Stapes extends from inner ear capsule to quadrate; quadrate small; quadrate-articular jaw joint
F: Early Gorgonopsians Eoarctops lower Upper Permian
Slightly larger than eye socket; expanded posteriorly and medially (minimal); 20-25% of skull length
65-75% of jaw length is the den- tary; posterior end of dentary slightly expanded posteriorly and dorsally as incipient coronoid process
Large single canine; other teeth simple cones.
40% of jaw length; angular notch deepened into a cleft; reflected lamina large, broad, blade-like
Stapes extends from inner ear capsule to quadrate; quadrate- articular jaw joint
E: Eotitanosuchians Sphenacodon Lower Permian
Small; slightly smaller than eye socket; slightly expanded posteriorly and medially
65-75% of jaw length is the den- tary; posterodorsal edge rises broadly but slightly above tooth row
Large single canine; other teeth simple cones.
40% of jaw length; angular notch deepened into a cleft; reflected lamina large, broad, blade-like
Stapes extends from inner ear capsule to quadrate; quadrate- articular jaw joint
D: Late sphenacodonts Sphenacodon Upper Pennsylvanian
Small; smaller than eye socket; confined to one side of skull
65% of jaw length is the dentary; posterodorsal edge rises broadly but slightly above the tooth row
Enlarged incipient canines; other teeth simple cones
60% of jaw length; venntral edge of angular notched ("angular notch") offsetting a short pro- tusion (reflected lamina)
Stapes extends from inner ear capsule to quadrate; quadrate large and plate-like; quadrate- articular jaw joint
C: Early spenacodonts Haptodus Upper Pennsylvanian
Tiny; smaller than eye socket; confined to one side of skull
65-75% of jaw length is the den- tary; posterodorsal edge rises broadly but slightly above tooth row
Undifferentiated; slightly enlarged incipient canines just behind nares
70% of jaw length; ventral edge of angular with shallow indentation
Stapes extends from inner ear capsule to quadrate; quadrate- articular jaw joint
B: Early ophiacodonts Archaothyris upper Middle Pennsylvanian
Tiny; smaller than eye socket; confined to one side of skull
x
Undifferentiated; slightly enlarged incipient canines just behind nares
x
Stapes extends from inner ear capsule to quadrate; quadrate- articular jaw joint
65-75% of jaw length is the den- tary; posterodorsal edge rises broadly but slightly above tooth row
Undifferentiated; slightly enlarged incipient canines just behind nares
70% of jaw length; ventral edge of angular continuous
Stapes extends from inner ear capsule to quadrate; quadrate- articular jaw joint
Table 1: Morphology of synapsid reptiles and mammals (Note that Hylonomus is a protothyrid, not a synapsid). Data from references cited in text.
Modern reptiles and mammals are very distinctive, easily diagnosable, and do not intergrade. Reptiles are covered by scales, mammals by hair; reptiles are cold-blooded, mammals warm-blooded; reptiles do not suckle their young, mammals have mammary glands; reptiles have sprawling posture, mammals have upright posture. Most of these features are soft part anatomy or physiology that very rarely fossilize (although dinosaur skin impressions are known from Cretaceous sediments, and imprints of mammal hair are known from Eocene bats from Germany; Franzen, 1990). In the fossil record, we must look to skeletal features.
There are many skeletal features which allow us to distinguish the reptiles from the mammals (Carroll, 1988; Table 1, rows A, M). The single most important defining characteristic is the nature of the articulation of the lower jaw to the skull (Simpson, 1959). In reptiles, multiple bones comprise the lower jaw. A small bone at the posterior end of the lower jaw, the articular, articulates with the quadrate bone of the skull (Simpson, 1959; Carroll, 1988). In mammals, one large bone, the dentary, comprises the lower jaw. It articulates with the squamosal bone of the skull (Simpson, 1959; Carroll, 1988).
From comparative anatomy studies, it is certain that most of the bones of the reptiles and mammals are homologous (Crompton & Parker, 1978; Carroll, 1988). Of greatest importance, the middle ear bones of mammals (stapes, incus, malleus, and tympanic) are homologous with several of the skull and jaw bones of reptiles (stapes, quadrate, articular, and angular, respectively; Romer, 1956, p. 33-38, 1970a; Allin, 1975, 1986; Allin & Hopson, 1992; Crompton & Parker, 1978; Hopso n, 1987, 1994; Carroll, 1988). One group of reptiles, the synapsids (Subclass Synapsida), share with the mammals an additional homologous structure: the lateral temporal fenestra, which is an opening in the skull behind the eye socket at the triple junction between the squamosal, jugal , and post orbital bones (Broom, 1932; Frazetta, 1968; Kemp, 1982; Carroll, 1988). A band of bone composed of the jugal and the squamosal is adjacent to the lateral temporal fenestra (Broom, 1932; Kemp, 1982; Carroll, 1988). This is the cheek arch so characteristic of mammal skulls (Broom, 1932; Kemp, 1982; Carroll, 1988). Therefore, synapsids are commonly named the mammal-like reptiles.
The presence of diagnosable morphologic differences between reptiles (including the oldest reptiles and the oldest synapsids) and mammals distinguishes them as distinct taxa. This allows us to test evolution by looking for transitional forms between the two. Because many of the bones are homologous, we should find evidence illustrating how these bones were modified over time to become the new bones. Furthermore, these morphologic changes should happen in parallel and in geochronologic succession.
Synapsid reptiles inhabited Pangea from the Middle Pennsylvanian through the Early Jurassic (Kemp, 1982, 1985; Sloan, 1983; Carroll, 1988; Hopson, 1969, 1987, 1994; Hopson & Crompton, 1969; Hotton, et al., 1986; Crompton & Jenkins, 1973; Sidor & Hopson, 1998; Romer & Price, 1940; Broom, 1932; Boonstra, 1963, 1969, 1971; Tchudinov, 1983; Olson, 1944; Tatarinov, 1974; Vyushkov, 1955; Efremov, 1954). From the Early Permian through the Early Triassic, they were the largest and most abundant land animals (Sloan, 1983; Colbert, 1965). Though much less well known to the general public than dinosaurs, one of the cereal box dinosaurs, Dimetrodon (the sail-backed reptile), is a synapsid, not a dinosaur (Romer & Price, 1940; Carroll, 1988). The oldest mammals are Late Triassic (Kemp, 1982; Carroll, 1988). Below is a discussion of the geochronologic succession linking synapsids and mammals. The oldest reptiles (named protorothyrids; Carroll, 1964, 1988, p. 192-199) are from the lower Middle Pennsylvanian, and the oldest synapsids (Reisz, 1972) are from the upper Middle Pennsylvanian, both of Nova Scotia. Upper Pennsylvanian and Lower Permian forms are known primarily from the midcontinent and Permian Basin region of the United States (Romer & Price, 1940; Currie, 1977, 1979; Kemp, 1982; Sloan, 1983). The basal Upper Permian forms are known from Russia (Tchudinov, 1960, 1983; Efremov, 1954; Olson, 1962; Sigogneau & Tchudinov, 1972; Ivakhnenko et al., 1997). Most of the Upper Permian and Lower Triassic succession is known from southern Africa, especially the Great Karoo of South Africa (Broom, 1932; Boonstra, 1963, 1969, 1971; Hopson & Kitching, 1972; Kemp, 1982; Sloan, 1983). The Middle Triassic forms are from South America (Romer, 1969a, 1969b, 1970b, 1973; Romer & Lewis, 1973; Bonaparte & Barbarena, 1975), and the Upper Triassic and Lower Jurassic mammals are known from Eurasia (Kermack, Mussett, & Rigney, 1973, 1981; Kemp, 1982). Subsequent Mesozoic mammals are known from all over the world (Simpson, 1928; Lillegraven et al., 1979).
When placed in proper geochronologic succession, the synapsids naturally form a succession of taxa (genera and families) that progressively become more mammal-like and less reptile-like (Kemp, 1982, 1985; Sloan, 1983; Sidor & Hopson, 1998; Hopson, 1987, 1994). Morphologic changes, summarized in Table 1 and Figure 1, affect the entire skeletal anatomy of these animals, but are most clearly displayed in their skulls.
The lateral temporal fenestra increased in size from a tiny opening smaller than the eye socket to a giant opening occupying nearly half the length of the skull. Ultimately, it merged with the eye socket, thus producing the full development of the cheek arch so characteristic of mammals (Broom, 1932; Frazetta, 1968; Kemp, 1982; Sloan, 1983; Hopson, 1987, 1994; Carroll, 1988).
Successively, the relative proportion of the lower jaw comprised of the dentary bone (teeth-bearing bone) gradually increased until the entire lower jaw consisted of the dentary (Kemp, 1982; Sloan, 1983; Carroll, 1988; Hopson, 1987, 1994). In Pennsylvanian and Lower and basal Upper Permian synapsids, the postero-dorsal edge of the lower jaw rose broadly but only slightly above the level of the tooth row (Romer & Price, 1940; Currie, 1977, 1979; Ivakhnenko et al., 1997; Tchudinov, 1960, 1983; Efremov, 1954; Olson, 1962; Sigogneau & Tchudinov, 1972; Hopson, 1987, 1994). In succeeding forms, the posterior part of the dentary expanded dorsally and posteriorly as a blade-like process, and progressively became larger (Broom, 1932; Boonstra, 1963, 1969, 1971; Sigogneau, 1970; Brink, 1963; Kemp, 1979; Hopson, 1987, 1994), forming the coronoid process (Parrington, 1946; Fourie, 1974; Romer, 1969b, 1970b, 1973; Hopson, 1987, 1994) to which the mammalian-type jaw musculature is attached (Barghusen, 1968; Bramble, 1978; Crompton, 1972; Crompton & Parker, 1978; Kemp, 1982; Sloan, 1983; Carroll, 1988). Concomitantly, the post-dentary bones progressively reduced in size (Allin, 1975; Crompton, 1972; Crompton & Parker, 1978; Kemp, 1982; Sloan, 1983; Carroll, 1988; Hopson, 1987, 1994).
Beginning with the Upper Pennsylvanian sphenacodonts, a notch developed in the angular bone that offsets a projection, the reflected lamina (Allin, 1975; Allin & Hopson, 1992; Hopson, 1987, 1994; Romer & Price, 1940; Currie, 1977, 1979; Kemp, 1982; Sloan, 1983; Carroll, 1988). The reflected lamina first became a large blade-like flange (Allin, 1975; Allin & Hopson, 1992; Hopson, 1987, 1994; Ivakhnenko et al., 1997; Tchudinov, 1960, 1983; Efremov, 1954; Olson, 1962; Sigogneau & Tchudinov, 1972; Broom, 1932; Sigogneau, 1970; Boonstra, 1963, 1969, 1971), and then was progressively reduced to a delicate horseshoe-shaped bone (Allin, 1975; Allin & Hopson, 1992; Hopson, 1987, 1994; Brink, 1963; Parrington, 1946; Fourie, 1974; Romer, 1969b, 1970b, 1973; Kermack, Mussett, & Rigney, 1973, 1981; Kemp, 1979, 1982; Sloan, 1983; Carroll, 1988).
Simultaneously, the quadrate progressively decreased in size (Allin, 1975; Allin & Hopson, 1992; Hopson, 1987, 1994; Kemp, 1982; Sloan, 1983; Carroll, 1988). The articular did not decrease in size much, being small initially, but developed a downward-pointing prong (Allin, 1975; Allin & Hopson, 1992; Hopson, 1987, 1994; Kemp, 1982; Sloan, 1983; Carroll, 1988). In the synapsids, the lower jaw was hinged to the skull by the articular and quadrate bones (Crompton, 1972; Crompton & Parker, 1978; Allin, 1975; Allin & Hopson, 1992; Hopson, 1987, 1994). Thus they are classified as reptiles (Simpson, 1959; Kemp, 1982; Sloan, 1983; Carroll, 1988). As the quadrate and articular became smaller, they were relieved of their solid suture to the dentary and skull (Crompton, 1972; Allin, 1975, 1986; Allin & Hopson, 1992; Hopson, 1987, 1994; Crompton & Parker, 1978; Kemp, 1982; Sloan, 1983; Carroll, 1988). A projection of the dentary extended posteriorly and made contact with the squamosal. Morganucodon possessed the mammalian dentary-squamosal jaw joint adjacent to the reptilian articular-quadrate jaw joint (Kermack, Mussett, & Rigney, 1973, 1981; Carroll, 1988). It is classified as the first mammal, but it is a perfect intermediate. Now that a new jaw joint was established, the quadrate and articular were subsequently relieved of that function (Crompton, 1972; Allin, 1975, 1986; Allin & Hopson, 1992; Hopson, 1987, 1994; Crompton & Parker, 1978; Kemp, 1982; Sloan, 1983; Carroll, 1988). Ultimately, in Middle and Upper Jurassic mammals, the tiny quadrate, articular, and ring-like angular migrated as a unit to the middle ear where they joined the stapes and became the incus, malleus, and tympanic bones (Allin, 197 5, 1986; Allin & Hopson, 1992; Hopson, 1987, 1994; Kemp, 1982; Sloan, 1983; Carroll, 1988).
Progressively, the teeth became differentiated. The large canines developed first, followed by the development of multicusped cheek teeth, reduced tooth replacement (Osborn & Crompton, 1973; Crompton & Parker, 1978), and finally full y differentiated incisors, canines, premolars, and molars with one tooth replacement during life (Kemp, 1982; Hopson, 1994).
Many other morphologic changes are documented in the fossil record. These demonstrate the morphologic and geochronologic succession from sprawling limb posture to upright limb posture of mammals (Jenkins, 1971; Romer & Lewis, 197 3; Kemp, 1982; Carroll, 1988; Hopson, 1994). As Jenkins (1971, p. 210) stated, In details of morphology and function, the cynodont post-cranial skeleton should be regarded as neither reptilian nor mammalian but as transitional between the two classes . Other changes have been adequately summarized elsewhere (Kemp, 1982; Sloan, 1983; Carroll, 1988; Hopson, 1994). Obviously, fundamental physiologic changes must have taken place as well, many of which are not directly preserved in the fossil record, though some can be inferred from the skeletal anatomy (Findlay, 1968; Kemp, 1982; Sloan, 1983, Carroll, 1988; Hopson, 1994).
This is well documented in the fossil record by a massive volume of incontrovertible data that cannot be explained away. Such large-scale, progressive, continuous, gradual, and geochronologically successive morphologic change (Sidor & Hopson, 1998) is descent with modification, and provides compelling evidence for evolution on a grand scale.
Is ID de wetenschappelijke (sic) uitleg voor creationiisten 150jaar Darwinisme... De evolutietheorie gewikt en gewogen David sorensen en revolutietheorie lalala DE VERRIJZENIS VAN DE JONGE VERDRONKEN KOE » Reageer (21) ID TOVERDOOS OVER SLECHTE ID-EETJES ,RESISTENTIE & TOVERDOCTORS c) OEC d) YEC
Een oeroud spoor Denisova : ZUID SIBERIË DENISOVA - mens
DMANISI AAP OF MENS DE OUDSTE ? » Reageer (1)Nakalipithecus nakayamai (<) Alweer eentje ? ..... De tand des tijds / Heidelberg-mens DE HEUPEN VAN EVA CASABLANCA MAN /erectus Hobbit is aparte soort ? » Reageer (2) NOG EEN BENDE BIJTERS POLONAISE met neanderthaler TANDEN UIT DE QESEM GROT Neanderthaler genoom Mitochondriale Genenkaart van Neanderthaler Xuchang mens
b)Biologie 1.-(EVOLUTIE ) NAS / IM Document 2008 (2) NAS / IM Document 2008 ( 1)
Cambrium & precambrium AVALON GABONESE chips
Evolutie in actie Opmerkelijk snelle adaptaties bij kroatische ruine-hagedissen:
EEN PLUIM VOOR CHINA Geef eens een pootje Pluimgewicht KLEIN DUIMPJE & DE REUS &VEREN Opnieuw gevederde Maniraptor Similicaudipteryx. Aerosteon riocoloradensis
Krokodillen
PAKASUCHUS KAPILIMAI Prestosuchus chiniquensis Terug naar zee viseter
Eritherium azzouzorum LJOEBA Darwinius masillae VLIEGENDE KATTEN ? Vleermuizen ALWEER EEN BELANGRIJK STUK UIT DE LEGPUZZEL Zee-zoogdieren ; Van de wal in de visgronden Indohyus / walvisevolutie Zeehonden-evolutie
*Blauwe en groene ( =nederlandse)teksten zijn meestal aanklikbare links *Engelse Wikipedia teksten verwijzen in de linkerkolom naar verschillende niet-engelse versies van het wikiartikel *Blauwe teksten tusssen "" , zijn voornamelijk ( gedeeltelijke)citaten afkomstig van mensen met andere meningen
Evolutie / Charles Darwin aan de basis: SELECTIE NATUURLIJK DE LEVENSBOOM UITSTERVEN DEEP TIME Biogeografie SEKSUELE SELECTIE Al jaren lang samen CO-EVOLUTIE De economie van de natuur Geleidelijke veranderingen
Unintelligent design:
IP Theorie Ons gebit , De appendix, Geboorte, De weg van het zaad , Nervus Laryngeus Recurrens, DNA , 75 mistakes , Het boek
Unintelligent design (1): Rechtop lopen Tomaso over het oog , oogspieren , Unintelligent Design (5): ZWEETVOETEN Unintelligent design (6): Vitamine C Unintelligent design (8): Kuitspieren
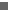



![[nature06277-f3.2.jpg]](http://2.bp.blogspot.com/_m6sVhskT_Fs/Ss98AEugXRI/AAAAAAAAEPI/8dy9_gADUME/s1600/nature06277-f3.2.jpg)

















)






![[Fig. 1a]](http://www.gcssepm.org/images/fossil_a.gif)
![[Fig. 1b]](http://www.gcssepm.org/images/fossil_b.gif)






 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}