Dit blog is een onderdeel van EVODISKU WAT IS DE BEDOELING EN WAAR STAAT DIT BLOG VOOR
****Wie meent dat alles inmiddels wel over de evolutietheorie gezegd is en dat de discussie gesloten kan worden, ziet over het hoofd dat de wetenschap niet stil staat.
***Wie meent dat inmiddels het creationisme definitief het pleit heeft verloren en dat de discussie gesloten kan worden , ziet over het hoofd dat het "creationisme" is geevolueerd ( en zal evolueren ) in nieuwere mimicrytische vormen( meme-complexen ) zoals bijvoorbeeld het ID(C)
***Dit blog is speciaal opgezet om de aktualiteit binnen het evolutie-creationisme debat te volgen en van kommentaren te voorzien ... waartoe de lezers zijn uitgenodigd bij te dragen ...
Let echter wél op het volgende :
"Je bent een rund als je hier met religie stunt " ....
Resten van mensachtigen van 4,4 miljoen jaar oud, waaronder een onvolledig skelet met schedel, zijn 17 jaar na de ontdekking eindelijk wetenschappelijk beschreven.
Wetenschappers zijn na een zeventienjarige studie van oeroude Oost-Afrikaanse skeletten tot de conclusie gekomen dat het om een oude voorloper van de moderne mens( en mogelijk ook de australopithecinae ?) gaat.
Ardi, of de ardipithecus ramidus, zwierf 4,4 miljoen jaar geleden rond in de bossen van Noordoost-Ethiopië
Excavating Ardi http://www.nytimes.com/slideshow/2009/10/01/science/100109_Fossil_index.html Excavating Ardi Ze was 1m20 groot, maar zeker geen lichtgewicht; haar gespierde lichaam woog ongeveer 50 kilogram. Ze klauterde met gemak in bomen dankzij haar lange armen, enorme handen en grijptenen, maar "Ardi" kon ook gewoon rechtop lopen. Ze is daarmee de eerste menselijke voorloper die op twee benen liep.
Door haar vacht en vooruitstekend gezicht moet Ardi er meer als een aap dan als een mens uit gezien hebben. Maar de "mozaik" aan kenmerken die Ardi in het verhaal van de oorsprong van de mens plaatst , tekent zich nu veel scherper af, na een uitgebreid onderzoek van 110 fragmenten van gefossiliseerde beenderen afkomstig van de soort waartoe ze behoorde.
Het is nu duidelijk dat ardipithecus ramidus onmiskenbaar een lid van een geslacht ( genus ) in een lange afstamming die leidt tot de homo sapiens. Die lijn start bij de laatste gemeenschappelijke soort uit een genus dat we delen met de chimpansees, onze (huidige ) dichtste verwanten, waarvan (het homologe ) DNA voor 95 a 96 procent met dat van ons overeenstemt.
Wetenschappers hebben ook vastgesteld dat op basis van het onvolledig skelet van een vrouwelijke ardipithecus Ardi is haar koosnaam de oudste, meest volledige verzameling ( van verschillende specimen van die soort afkomstige )gefossiliseerde overblijfselen ( en model- reconstructie ) is van de vele voorlopers uit die oer- tijd en die voortkwamen sinds en uit de gemeenschappelijke voorloper( Last Common Ancestor ) , een tot dusver (nog fossiel) onbekende soort die waarschijnlijk 6 tot 7 miljoen jaar geleden op aarde rondliep.
Zeventien jaar lang bestudeerden wetenschappers elk aspect van de anatomie en de habitat van de ardipithecus. Deze presenteerden ze het resultaat van hun onderzoek, in de vorm van elf aparte wetenschappelijke bijdragen in het magazine Science.
"Deze fossielen verzameling ( en de huidige studies in Sciencemag ) is een van de belangrijkste ontdekkingen/ rapporten voor de studie van de menselijke ontwikkeling", aldus David Pilbeam, curator in het Peabody Museum of Archeology and Ethnology van de Universiteit Harvard. "(De reconstructie ) is vrij compleet, want hoofd, handen, voeten en enkele andere belangrijke delen zijn overgebleven(van de eerste vondst ) Het vertegenwoordigt een geslacht dat mogelijk voorafging aan de Australopithecus, het geslacht dat aan ons geslacht Homo voorafging."
De foto op de cover Van Science Geplette schedel , een aantal kiezen en tanden , twee armen, het bekken, een been en twee voeten .... http://johnhawks.net/weblog/fossils/ardipithecus/limb-proportions-photo-note-2009.html " .....The composite photo included in the main paper by White and colleagues (2009) is very poorly scaled. For example, the measurements given by Lovejoy and colleagues (2009) make the radius 95 percent the length of the femur, but the photo renders the radius quite a bit longer than the tibia:This has the effect of making the photo look much more like the proportions of a chimpanzee or gorilla (or Oreopithecus...) than is the case from the skeletal measurements.
Although I will say that the Matternes reconstruction has much more slender-looking hindlimb bones than the photos. Since one of the interesting aspects of Ardi is the large reconstructed mass (51 kg), I find the very gracile long bone reconstruction misleading, also.
Het vakblad pakt groot uit met het belangrijke nieuws over " ardi " , dat feitelijk een gedegen reconstructie is (er is 17 jaar aan gewerkt !!! en er grepen al die jaren vervolgonderzoeken en preparaties van het fossiel materiaal , plaats ) van een vroege mensachtige(4,4 miljoen jaar oud) waarvan resten in 1992 en 94 -95 zijn gevonden, en daardoor onze ( fossiel )best gedocumenteerde (tot nu toe ) bekende (zeer waarschijnlijk) rechtoplopende voorouder. Sciencemag komt met een speciale editie met liefst 11 wetenschappelijke artikelen die allen een ander aspect van de vondst belichten.
Kloof gedicht ? Voor het onderzoek werden verdere fossielen gezocht en gevonden op het terrein, die vervolgens in laboratoria geanalyseerd werden. Daaruit blijkt nu dat de vroege menselijke voorouder de kloof overbrugt tussen de in bomen levende, van ver verwante primaten en onze op twee benen lopende voorouders die het bos als habitat hadden ingeruild voor het grasland van de savanne.
Een van de verrassendste bevindingen is dat deze voorouders niet op hun knokkels steunden, zoals altijd werd aangenomen. Ardi kon trouwens niet rondlopen op haar "knokels " : wegens de té flexibele en voor dat doel té tere pols Soepele polsen , sterke opponeerbare duimen , flexible vingers en handpalmen met stevige grip : lijken te zijn aangepast aan het grijpen van stenen en stokken ..Ze lijken voorbestemd om , miljoenen jaren later , vuistbijlen en andere stenen werktuigen te hanteren en te maken ...en wat later instrumenten te bedienen ....
Het is nu duidelijk dat de chimpansees van vandaag even drastisch afwijken van onze laatste gemeenschappelijke voorloper als de mens van vandaag.
"Charles Darwin zei heel verstandige dingen over die kwestie. Hij zei dat we voorzichtig moeten zijn. De enige manier om iets te leren over die gemeenschappelijke voorouder is ernaar op zoek gaan", zegt professor Tim White van de University of California Berkeley, een van de leiders van het onderzoek. "Wel, vondsten van 4,4 miljoen jaar geleden komen dicht in de buurt. Zoals Darwin al stelde, zijn de mens en de aap onafhankelijk van elkaar geëvolueerd sinds hun pad scheidde, sinds de laatste gemeenschappelijke voorouder."
Nota : ...Wat Darwin werkelijk heeft geschreven aangaande dit alles zou ik wel eens willen weten en/of duidelijker gerefereerd willen zien ....of legt men hier Darwin simpelweg woorden in de mond ? Zie hierover vooral john Hawks http://johnhawks.net/weblog/reviews/tv/discovering-ardi-release-2009.html Het verhaal van de ardipithecus begon in december 1992, toen een student van professor White, Gen Suwa, in de buurt van het dorp Aramis in Noordoost-Ethiopië , weerkaatst zonlicht bemerkte op het gladde oppervlak van een maaltand (=kies ) van een mensachtige.... de lijn die vertrok van de laatste gemeenschappelijke voorouder.
Een paar jaar later hadden wetenschappers een rijke collectie gefossiliseerde beenderen verzameld van de ardipithecus ramidus. Datering wees uit dat de beenderen 4,4 miljoen jaar oud waren, meer dan een miljoen jaar ouder dan de oudste afstammeling tot dan toe, een soort die bekendstaat als de australapithecus afarensis, Ardi had net zoals Lucy hersenen, niet veel groter dan die van een chimpansee en ongeveer een derde van die van de moderne mens.
De schedelbasis van Ardi, de afstand tussen de achter- en voorkant van de schedel, is kort, wat aangeeft dat het hoofd op de ruggengraat rustte, zoals bij andere rechtoplopers, en geen hoek maakt met de ruggengraat, zoals bij apen.
In tegenstelling tot Lucy vertoonde Ardi wel nog een paar anatomische kenmerken in voeten en armen die erop wijzen dat ze ook in bomen klom. Met haar handen kon ze objecten vastgrijpen. Wellicht was dat een essentiële vaardigheid die primaten in staat stelde om zo enorm intelligent te worden. Zo konden ze dingen oprapen, ze manipuleren en in het geval van mensen en chimpansees ook als instrumenten gebruiken. Maar het zou nog een paar miljoen jaar duren voor de afstammelingen van Ardi de grote hersenen en de hogere intelligentie ontwikkelden die de mens onderscheiden van de rest van het dierenrijk. Het is nu ook duidelijk dat de toename van het hersenvolume pas plaatsgreep lang nadat de primaten op twee benen waren gaan lopen.
® The Independent /Publicatie datum : 2009-10-03 (De Morgen )
Ondanks de dagelijkse zoektocht naar fossielen van onze vroegste voorouders, hebben paleontologen nog maar een zestal (meestal niet volledige) skeletten van oude mensachtigen kunnen vinden. Het oudste dat in een wetenschappelijke publicatie is beschreven, is dat van Lucy, de Australopithecus afarensis van 3,2 miljoen jaar oud die in 1974 werd gevonden in de Afardriehoek in Ethiopië. Vlakbij, bij het dorpje Aramis, vonden wetenschappers in 1992 andere fossiele resten.
Reconstructie van het skelet Foto's : J.H. Maternes Ardipithecus was 120 cm lang en woog 50 kilo. Best zwaar en groot voor zon nog altijd aapachtige mensachtige.
-De in 1992 gevonden geplette schedel en het gedeeltelijke skelet waren dat van een vrouwelijke mensachtige, die de onderzoekers Ardipithecus ramidus noemen.
-Het hoogtepunt in de zoektocht en reconstructieproces lag in 1994-95, toen het team van Tim White onder een verhoging in het terrein stukjes van een bekken, een been, een voet en een schedel vonden, kortom een onvolledig skelet.
Ardi heeft veel meer dan Lucy sterkte aapaachtige kenmerken en had de bouw en de enorme grijphanden en voeten voor een (gedeeltelijk?) leven in de bomen.
Ardi's handen en polsen vertonen zowel primitieve kenmerken als modernere. -Een aapachtige of mensachtige hand ? ( verschil is te vinden in de vergelijking van het belangrijkste handwortelbotje)
Two views of the left hand of Ar. ramidus showing primitive features absent in specialized apes. (A) Short metacarpals; (B) lack of knuckle-walking grooves; (C) extended joint surface on fifth digit; (D) thumb more robust than in apes; (E) insertion gable for long flexor tendon (sometimes absent in apes); (F) hamate allows palm to flex; (G) simple wrist joints; (H) capitate head promotes strong palm flexion. Inset: lateral view of capitates of Pan, Ar. ramidus, and human (left to right). Dashed lines reflect a more palmar capitate head location for Ar. ramidus and humans, which allows a more flexible wrist in hominids. http://www.sciencemag.org/cgi/content/full/326/5949/70/DC2
Haar handpalmen waren kort en haar vingers flexibel, ze kon zich niettemin in een boom voortbewegen .
Ze had een unieke opponeerbare dikke teen . Tegelijk staan haar bekken en het achterhoofdsgat zodanig dat ze vrijwel zeker rechtop kon lopen. Mogelijk liep ze ook regelmatig staand door de bomen,( zoals bijvoorbeeld Gibbons soms doen ) over de takken, denken de ontdekkers onder leiding van de bekende paleontoloog Tim White. Mogelijk is ardi een overgangsvorm tussen een leefgedrag in de bomen door van boom naar boom te slingeren en rechtoplopen. De onderzoekers sluiten wel uit dat Ardi kon knokkellopen, zoals moderne chimpansees doen
* Ardi is een mensachtige die kan gelden als een mogelijk voorouder of een nauwe verwante aan een voorouder van australopithecus en latere homini . Ardi leefde ongeveer twee miljoen jaar na de geschatte laatste gemeenschappelijke voorouder van mens en apen, die rond de zes a zeven miljoen jaar(= de zogenaamde "split " = is berekend /geschat /hypothetisch voorspeld , volgens de 'moleculaire' klok ) geleden in Afrika voorkwam. Door de vulkanische lagen boven en onder het skelet te dateren kon de geoloog Giday WoldeGabriel bepalen dat eerstgevonden Fossiel 4,4 miljoen jaar oud is. Dat de fossiele resten miljoenen jaren oud waren, bleek ook uit de staat van het bot: bij de minste aanraking kruimelde het in stukken uiteen. De onderzoekers besloten niet alles ter plaatse met borstel en penseel uit de grond te halen, maar ze groeven grote blokken sediment uit en transporteerden die naar labs in de Ethiopische hoofdstad Addis Abeba, naar Tokyo en naar Ohio. Tim White deed er jaren over om in het Nationaal Museum van Addis Abeba all de kleigrond van de fossielen te verwijderen. Gen Suwa, de Japanse student die in 1992 de tand vond, puzzelde de 65 stukjes van de schedel in zijn lab in Tokyo virtueel weer in elkaar. Een andere medewerker van White, Owen Lovejoy deed hetzelfde voor het bekken in zijn lab in Ohio. Suwa was pas tevreden met de 10de reconstructie, die in maart dit jaar klaar was. Lovejoy had 14 pogingen nodig.(2-)
Dat Ardi ook rechtop kon lopen blijkt uit de manier waarop haar bilspieren op haar bekken en heupen lagen en waarmee ze zich in evenwicht kon houden. Daarnaast waren haar voeten stug genoeg om op te lopen, hoewel er ook nog steeds een duim aanwezig was om mee te kunnen klimmen.
Foot skeleton of Ar. ramidus (bottom; reconstruction based on computed tomography rendering shown) lacked many features that have evolved for advanced vertical climbing and suspension in extant chimpanzees (pan, top left). Chimpanzees have a highly flexible midfoot and other adaptations that improve their ability to grasp substrates. These are absent in Ar. ramidus. Credits: Reconstruction; copyright J. H. Matternes; chimpanzee climbing, J. DeSilva; bonobo and human feet, S. Ingham. http://www.sciencemag.org/cgi/content/full/326/5949/72/DC2
Over het rechtoplopen is niet iedereen het eens, maar feit is dat haar bekken even breed is als dat van Lucy( een veel jongere afstammeling of verwant ? ) , die ook rechtop kon wandelen
The Ar. ramidus pelvis has a mosaic of characters for both bipedality and climbing. Left to right: Human, Au. afarensis (Lucy), Ar. ramidus, Pan (chimpanzee). The ischial surface is angled near its midpoint to face upward in Lucy and the human (blue double arrows), showing that their hamstrings have undergone transformation for advanced bipedality, whereas they are primitive in the chimpanzee and Ar. ramidus (blue arrows). All three hominid ilia are vertically short and horizontally broad, forming a greater sciatic notch (white arrows) that is absent in Pan. A novel growth site [the anterior inferior iliac spine (yellow arrows)] is also lacking in Pan. http://www.sciencemag.org/cgi/content/full/326/5949/71/DC2.
3D-reconstructie van de schedel van Ardipithecus ramidus, een 4,4 miljoen jaar oude voorganger van de mens. (Illustratie Science) Haar hersenen hadden de omvang van die van een chimpansee.
Digital representations of the Ar. ramidus cranium and mandible. (A to D) The ARA-VP-6/500 and downscaled ARA-VP-1/500 composite reconstruction in inferior, superior, lateral, and anterior views (in Frankfurt horizontal orientation). (E) Individual pieces of the digital reconstruction in different colors. Note the steep clivus plane intersecting the cranial vault on the frontal squama (as in Sts 5 and not apes). (F and G) Lateral and superior views of the ARA-VP-1/401 mandible (cast). (H and I) Lateral and superior views of the ARA-VP-6/500 left mandibular corpus with dentition. http://scienceblogs.com/pharyngula/
*Mannetjes en vrouwtjes missen de zware hoektanden ( niet zoals chimps, maar net net zoals wij ) Het suggereert dat er GEEN uitgesproken sexueel dimorphisme was zoals bij chimpansees en gorilla's ( en het genus paranthropus ) .... Dat suggereert ook( althans dat is een speculatie van C. Owen Lovejoy http://johnhawks.net/node/2246 * ) dat er geen sterke man-man competie was( zoals bij chimps ) bij deze verre voorouder Maar het is nu eerst en vooral zoeken naar een mannelijke schedel zonder kam ; die deze vermoede eigenschap zou kunnen bevestigen ,en vooraleer dergelijke zwakke maar verregaande speculaties als "zeer waarschijnlijk " voor te stellen *het tandemail (dunner dan bij Lucy ,maar dikker dan datvan een chimp) wijzen op een omnivoor dieet :fruit , planten , insecten en kleine zoogdieren . Ardi en Lucy bezaten een andersoortig gebit in de details ; Lucy had grotere maalkiezen die eigenlijk geschikter waren op de savanne * Ardi kan natuurlijk ook een voorouder zijn in de basis van de australopithecinae-lijn
Dentitions from human (left), Ar. ramidus (middle), and chimpanzee (right), all males. Below are corresponding samples of the maxillary first molar in each. Red, thicker enamel (~2 mm); blue, thinner enamel (~0.5 mm). Contour lines map the topography of the crown and chewing surfaces. http://www.sciencemag.org/cgi/content/full/326/5949/69/DC2
Ardi leek niet echt op chimpansee De basis van de schedel toont aan dat Ardi niet met het hoofd voor haar ruggegraat liep, zoals bij viervoetige apen. Ook haar tanden lijken niet op die van moderne chimpansees geen scherpe bovenste hoektanden.
Volgens de onderzoekers toont dit aan dat de Ardipithecus ramidus helemaal niet op een chimpansee leek, en zich ook niet zo gedroeg.
Alan Walker, antropoloog aan Pennsylvania State University, zegt in Science dat uit de vondst blijkt dat we aan de hand van de anatomie van de aan ons verwante diersoorten (zoals de chimpansee), niet altijd kunnen leren over de lichaamsbouw en het gedrag van onze voorouders.
Het ziet ernaar uit dat de laatste gemeenschappelijke voorouder van mens en chimpansee veel minder op de moderne chimpansee leek dan we totnogtoe dachten.(1) "Ardipithecus is not a chimp. It's not a human. It shows us what we used to be." (Professor Tim White University of California Berkeley, one of the leaders of the research.)
Het skelet vertoont wél aapachtige kenmerken die niet voorkomen in moderne chimpansees en gorilla's. Dat betekent, schrijven ze nog in Science, dat ook de apen in Afrika nog een ingewikkelde ontwikkeling hebben doorgemaakt nadat ze in de evolutie hun eigen weg gingen (1)
Onze voorlopig oudste voorouder had meer gemeen met een mens dan met een aap en leefde in dichte regenwouden.
De nieuwe mensachtige heet Ardipithecus ramidus en haar anatomie ondergraaft enkele wijdverspreide inzichten in de evolutie van de mens. Ardi zoals ze de voorloper van Lucy noemen kon volgens het internationaal team van onderzoekers rechtop lopen, maar ook in bomen leven. Toch leek ze allerminst op een moderne chimpansee: ze liep niet op de knokkels, had een rechtopstaand hoofd en een rechte rug.
Professor White grapte ooit tegen een collega dat als je wilt weten hoe Ardi zich voortbewoog, dat je dan naar de bar in Star Wars moet gaan.
"De ardipithecus is geen chimpansee en geen mens. Het is wat wij ooit waren. Het is een mozaïekschepsel dat noch chimpansee noch mens is."
Zijn collega professor Owen Lovejoy van Kent State University voegt ( er nogal ongelukkig ) aan toe: "De nieuwe anatomie die we beschrijven in deze artikels wijzigt op fundamentele wijze onze kennis over de oorsprong van de mens en de vroege evolutie."
Hoewel laatste gemeenschappelijke voorouder van mens en chimpansee 6 miljoen jaar geleden leefde, leken de voorouders van de mens dus helemaal niet op moderne chimpansees. "Ardi is niet datgene wat men zou hebben voorspeld ... " zei antropoloog Bernard Wood (George Washington University.)
Een ongelukkig zinnetje waar natuurlijk door allerlei creationisten in de toekomst zal worden ( en nu al wordt geciteerd uit de context) naar verwezen Het is gebaseerd op een soort misvatting / en verdraaing gebaseerd op een gebrek aan kennis over wat voorspelbaar en niet voorspelbaar is in de paleontologie en o.a. door reeds hieronder door Bart klink en vooral Gerdien De Jong afdoende is aangekaart
-Ze was niet een tussenvorm tussen een mens en een chimpansee, maar anders.... BK: Natuurlijk niet, er is ook niemand met verstand van evolutie die dat ooit beweerd heeft.
-Zij liep rechtop, en niet zoals chimpansees op handen en voeten.? BK: Ardi liep rechtop, maar niet zo goed als wij dat kunnen en kon zeker niet goed rennen. Hetzelfde geldt ongeveer voor Lucy.
Dichtbegroeid regenwoud Sinds Lucy staat in de schoolboeken dat de mensachtigen rechtop begonnnen te lopen op grasland, en niet in bossen. Maar uit de analyse van fossiele planten- en dierenresten die in de buurt van het oude graf van Ardi werden gevonden, blijkt dat het gebied vroeger een dicht regenwoud was, met vele bomen waartussen Ardi soms eens slingerde van tak tot tak, en soms tussen de bomen op de grond gewoon rechtop wandelde.
-Ardi voldoet niet aan de verwachting dat een voorloper van de moderne mens op de open savanne leefde.? BK: De homininen liepen niet vanaf het prille begin op de open savanne, zoals blijkt uit bijvoorbeeld Orrorin. Gerdien de Jong : Het idee van de open savanne begint pas met Homo erectus: de eerste lange afstandsloper.
Chris verburgh Opvallend is dat de Ardipithecus zowel aanpassingen had om in bomen te leven, als om rechtop te lopen op de grond. Dat zou kunnen impliceren dat onze voorouders al rechtop liepen toen ze nog tussen en in de bomen leefden, en niet daarmee zijn begonnen toen Oost-Afrika bezaaid was met savannevlaktes. Eén van de populaire theorien die het rechtop lopen wil verklaren, ging er immers van uit dat onze voorouders rechtop begonnen te lopen omdat de bomen verdwenen om plaats te maken voor savannevlaktes en ze dan wel verplicht waren om op de grond te leven.
2.- Belangrijk Kommentaar van Gerdien De Jong comments #9 " .....Ik ben het volledig met Bart Klink eens dat een wezen met grofweg deze eigenschappen in deze tijd voorspelbaar was. Op grond van Sahelanthropus, Ororin en Lucy: rechtoplopend, met relatief lange armen.
*Je kunt voorspellen dat je - Homo erectus buiten Afrika op het vasteland van Eurazie zult vinden, gegeven dat Homo erectus om te beginnen op Java gevonden is, en alle eerdere hominiden in Afrika gevonden zijn. Ook kun je voorspellen uit welke tijdsperiode dat zou kunnen zijn: -na verschijnen in Afrika en voor opduiken op Java. Dat zijn allemaal vrij gemakkelijke voorspellingen: alleen maar samenhang in de gegevens gevraagd.
*Wat je niet kunt voorspellen is dat de oudste Homo erectus buiten Afrika in Georgie gevonden zou worden.
* Wat je wel kunt voorspellen is dat het de moeite waard is de Sahara en de Sahel (3)grondig voor fossielen af te zoeken als je op zoek wilt naar iets dat nog net een chimpansee voorouder kan zijn net als Arabie en Perzie voor een nog iets vroegere voorouder als ik de geografie van de Miocene mensapen goed heb onthouden.
*Er was niet te voorspellen dat er een Flores mens zou opduiken, of dat dat een Homo erectus vorm zou zijn. Wel was te voorspellen dat als er een Homo erectus op Flores zou hebben overleefd, die kleiner zou zijn dan de Homo erectus van grote eilanden en continenten.
*Niet voorspelbaar was de volgorde van veranderingen voordat er voldoende fossielen gevonden werden. Vandaar dat je in de oudere (intussen behoorlijk oudere) literatuur veelal vindt dat bipedalisme laat zal zijn.
*Wat duidelijk is, is dat voorspellingen uit de losse pols niet mogelijk zijn. Voorspellingen als: de samenhang van de fossielen vraagt (xx) zijn duidelijk wel mogelijk. Daaronder vallen Tiktaalik,(3) Anchiornis en Ardipithecus. Dit soort voorspellingen is natuurlijk niet te doen zonder een behoorlijk groot fossiel archief. . De Flores mens was niet voorspelbaar, omdat er geen fossielen waren waarbij de samenhang van de fossielen een Flores mens vroeg.
De wetenschappers die Ardi bestudeerden beweren echter NIET dat ze noodzakelijk of effectief evolueerde tot de australopithecinen en/of rechtstreeks tot de mensen van het geslacht homo leide ( dat is slechts één van de hypotheses ) Over Australopithecus & co (Tsjok45)<--(klik)
ook hier kan de ergvroege voorouderlijke oer-menselijke(homo) lijn zeer goed in allerlei aftakkingen zijn opgesplitst ( = een struik), waarbij de meesten zijn uitgestorven ( denk bijvoorbeeld ook ter vergelijking aan ( de veel later )optredende vertakkingen in het genus homo zelf--> Homo floresiensis en Neanderthaler, Australopithecinen zelfs ) Lucy's lijn kan bovendien zijn afgesplit van ardi's lijn lang voordat ardi leefde... Maar zelfs dan , zegt White te geloven dat zijn team een evolutionaire sequentie ( op het genus-niveau )heeft gedocumenteerd waaruit het genus homo uiteindelijk onstond( metof zonder de tussenstap Australopithecus ) Ardipithecus--->(mogelijk) dan Australopithecus, ----> Homo.
"Ardi vertelt ons toch meer dan Lucy We bezitten haar handen en voeten , een compleet beeld van haar leefomgeving , een complexer skelet, ouder , primitiever : Ardi toont ons hoe het de transformatie van gemeenschappelijke voorouder naar hominide kan zijn gebeurt ... " C. Owen Lovejoy
Three hypotheses of early hominin evolution. Top - That there was a straight line of hominin evolution, each "species" being the phase or grade of one true natural species. Middle - A single line of hominin evolution with a speciation event in the same area in which the earliest Australopithecus split from Ardipithecus ramidus. Bottom - Ardpithecus ramidus as one of the last members of a more archaic lineage which existed after a speciation event elsewhere in Africa that gave rise to the first Australopithecus. Published in this week's issue of Science.
(*)DezeC.Owen lovejoy is blijkbaar niet vies van het verkopen van nonsens en het doen van "sensationele "uitspraken ? http://johnhawks.net/node/2246 C. Owen Lovejoy, an anatomist at Kent State University and lead author of two of the journal reports, speculated that these hominids had a social system that involved less competition among males and that this suggested the beginning of pair bonding between males and females. Dr. Pilbeam disputed this conjecture, saying, This is a restatement of Owen Lovejoys ideas going back almost three decades, which I found unpersuasive then and still do. paleoanthropologist Andrew Hill of Yale University.was more blunt, calling Lovejoy's speculation "patent nonsense." http://www.latimes.com/news/nationworld/nation/la-sci-fossils2-2009oct02,0,3420742.story?track=rss De vondsten "are turning evolution on its head," zei Lovejoy( overigens een verdienstelijk en uitstekend wetenschapper die evengoed lang op zijn blote knieeen in het zand heeft gezeten tijdens ( ook deze ) opgravingen en expedities ) maar hij trekt wél de aandacht met zijn voorbarige miskleunen voor de camera's en de persmuskieten : Binnenkort staat ie in de kijkcijfers en misschien zelf eens op zijn hoofd kunstjes te maken en de aandacht verder op te eisen ?
http://sandwalk.blogspot.com/2009/10/good-science-bad-science-journalism.html Natuurlijk moeten ook Wetenschappers( Hier O.Lovejoy ) weer onvoorzichtige uitspraken doen , waarna journalisten fluks overgaan op hun geliefkoosde " conclusion jumping " en allerlei onzin gaan uitkramen omdat het o.a. zogenaamd"sensationeel" is het en geschikt voor de vluchtige oppervlakkige doorsnee lezer /kijker op zoek naar infotainment
(L.Moran )".....The press reports convey the message that, up until last week, scientists thought that humans evolved from chimps. Now, with the publication of the Ardi papers, scientists have to change the textbooks and teach that chimps evolved from humans.
Is that really what scientists want the general public to think? It makes them look very stupid and (therefore ) the public has a right to ask what "other false notions about evolution "are being taught in the universities.
Voor wie het nog niet wist * Mensen stammen niet af van apen * ( en het omgekeerde )Chimpansees stammen ook niet af van mensen Maar het ZIJN wél allebei (mens)apen
Er is gelukkig ook nog altijd betere journalistiek = Carl Zimmer , natuurlijk http://blogs.discovermagazine.com/loom/2009/10/01/ardipithecus-we-meet-at-last/ "...Just a Reminder: We Didnt Evolve From Chimpanzees Chimpanzees may be our closest living relatives, but that doesnt mean that our common ancestor with them looked precisely like a chimp. In fact, a lot of what makes a chimpanzee a chimpanzee evolved after our two lineages split roughly 7 million years ago. Ardipithecus offers strong evidence for the newness of chimps..."
http://scienceblogs.com/pharyngula/ Wat literalistische geloofsgekken en Creato's ( misschien )zouden kunnen zeggen ( volgens een ironische PZ Meyer ) ? : " ....This only shows that humans were created first, and all of these other hairy beasts the paleontologists are digging up are just degenerate spawn of the Fall....? "
1) Er zijn tot nu toe geen oude fossielen ( dicht bij de split ) uit de chimp-lijn bekend ... buiten een paar tanden ... Het is derhalve(nog)niet mogelijk vroege mensen en vroege chimps ( en hun gemeenschappelijke voorlopers) te vergelijken Creato's (2) Alvast één voorspelbare reactie : ..17 jaar (zogenaamd) onderzoek om leugens en just so stories , te fabriceren ? want Die atheisten en hun wetenschappelijke samenzweerders zijn tot alles in staat om de mensheid te bedriegen en te brain-washen ....
De stamboom van de mens staat opnieuw op zijn kop ? * er wordt helemaal niets gezegd over een "rechtstreekse voorouder" of over een "missing link " , tenzij in kranten * De mogelijke verwantschappen werden opgesteld als denkbare sequenties op het genus-niveau * Er IS ook helemaal geen stamboom(= struik ) van de hominem die op zijn kop wordt gezet door de eindelijk gepubliceerde beschrijving van Ardipithecus ramidus ( er is zelf nog een oudere verwant bekend ---> Ardipithecus Kabbada ? ) Integendeel zelfs : de mix aan eigenschappen die ardipithecus vertoont ( een mozaik)is datgene wat paleantropologen grofweg mochten verwachten van een fossiele voorouder van de "hominim" uit die tijd en op die plaats ...
Hoe gaan creationisten eigenlijk reageren ? Al even "voorzienbaar "als het bovenstaande : is deze opsomming van mogelijke argumenten ( en omdat de eerste reacties bij amerikaanse crea's zijn te verwachten is het uit enkele Engelstalige diskussies , overgenomen Bovendien zijn amerikaanse creationisten nog steeds de modellen van de crea's en inspiratiebron voor complotdenkers en anti-wetenschappers , wereldwijd ) * How will creationist handle this ? (Tsjok45)<--klik
Al jazeera & Ardi *volgens de Arabische nieuwsdienst zou verder onderzoek naar Ardipithecus ramidus aantonen dat de evolutietheorie verkeerd is. Volstrekte onzin natuurlijk, en opvallend is dat er op de Engelstalige site van Al Jazeera geen enkele link is naar het wereldschokkende artikel.
* Uiteraard konden ook sommige amerikaanse journalisten het niet laten om het publiek te misleiden en te schrijven dat de Ardipithecus vondsten Darwins evolutietheorie weerleggen. Chris Esparza van de Christian Living Examiner schrijft: "The idea of the missing link is that somewhere way back when, there was a primate who almost seemed to be half monkey and half human, proving that there was at some point an evolutionary split. A recent discovery in Ethiopia disproves that theory." *De Ardipithecus is nu net een prachtvoorbeeld van een 'tussensoort' (die eigenlijk zoals altijd een volwaardige soort op zich is). *Het is duidelijk dat Chris Esparza nooit de moeite heeft genomen éérst te weten wat de theorie precies zegt, en dàn pas te onderzoeken of het juist dan wel fout is. Meer in het bijzonder over hoe die tussensoort er uitziet zegt hij bepaald niet wat "the idea" was, maar integendeel wat een stel andere onwetenden hem hebben wijsgemaakt wat "the idea" was. Als je wil weten wat "the idea" werkelijk was kan je altijd Darwin zelf raadplegen - even voor het gemak aannemend dat Darwin ons kan vertellen wat de evolutietheorie zegt. Hier gaan we: I have found it difficult, when looking at any two species, to avoid picturing to myself, forms directly intermediate between them. But this is a wholly false view; we should always look for forms intermediate between each species and a common but unknown progenitor; and the progenitor will generally have differred in some respects from all its modified descendants. (Darwin, The Origin of species, hoofdstuk 9)
(3) Maar al te vaak wordt stilletjes vergeten dat de Paleontologie ook een geologische deel-wetenchap is -Van Tiktaalik en co ( er is namelijk een hele reeks gevonden ) werd voorspeld in welke geologische lagen , formaties en geologische ouderdomsperiode moest worden gezocht naar de overgansvormen tussen vissen- amfibieen ... -Net zo kan men wisselend succes in de prospectie van petroleumvoorraden en kolenlagen (gedeeltelijk) voorspellen *(naar Bart klink ) " Waarom gaan paleontologen dit soort mensachtige voorouderlijke fossielen gaan zoeken in Afrika, in Pliocene lagen? -Juist omdat ze voorspeld hebben dat die fossielen daar zitten! -Paleontologen doen niets anders in de praktijk. Ze weten waar ze moeten zoeken, juist op grond van voorspellingen. -of, is het toeval dat ze juist daar zijn gaan zoeken? " - dat geldt dus ook allemaal voor de voorspelling dat in de Sahel en Sahara ( ooit beboste waterrijke gebieden ) nog meer fossielen zullen worden gevonden die nog meer licht gaan werpen op de "split " Tot nu toe is daar o.a. de Sahelantropus Tchadensis , en Praeanthropus(?= Australopithecus ) bahrelghazali, gevonden Het is dus wachten op de definitieve beschijving van vooral de eerstgenoemde= Toumai
Reacties op bericht (20)
30-06-2015
john
Cambodia has approved the transfer of four refugees from Australia as part of a controversial agreement under which the south-east Asian nation has received almost $40 million in additional aid from Australia. Please visit online: http://visasolutions.com.au
30-06-2015 om 12:42
geschreven door john
john
These cameras can work round the clock to ensure that you can see your property any time you want. Visit this link: http://www.foscamwa.com.au/
30-06-2015 om 12:42
geschreven door john
john
What's Microsoft Workplace 365 and how could you assess whether it is befitting your company? This post explores a few of the pros as well as cons that will help you along the right path. Click here: http://www.iquest.com.au/
30-06-2015 om 12:41
geschreven door john
john
If you ever and your enterprise have undoubtedly implemented quite a few virtualized methods, you certainly realize the benefits of with the ability to manage a person's network's total capacity. But for the reason that resources will be distributed differently inside of a virtualized design, being qualified to properly accomplish when allocating resources might be a problem. Please check out my page for more information about : https://www.zettagrid.com/
30-06-2015 om 12:40
geschreven door john
john
Personal private server combines the top qualities involving dedicated along with shared hosts. It gives a superlative web hosting service solution which is not only accommodating and scalable, and also cost powerful. Here can be a few things that you can keep planned while selecting a first-rate VPS web hosting service solution. Visit our website: http://www.zetta.net.au/
-- one ping per second.If the Ocean Shield can box in the pingers, the search area will be narrowed to within "a couple of hundred meters of the target,"
09-04-2014 om 08:33
geschreven door Michael Kors Outlet Online
17-03-2014
ENGD
I really enjoy simply reading all of your weblogs. Simply wanted to inform you that you have people like me who appreciate your work. Definitely a great post. Hats off to you! The information that you have provided is very helpful.
I am very happy to be here because this is a very good site that provides lots of information about the topics covered in depth. Im glad to see that people are actually writing about this issue in such a smart way, showing us all different sides to it. Please keep it up. I cant wait to read whats next.
This is like my
fourth time stopping over your Blog. Normally, I do not make comments on
website, but I have to mention that this post really pushed me to do so.
Really great post . Buy Codeine
19-12-2013 om 22:51
geschreven door john
boxer
Sometimes we are very concerned about our health, do not understand what
to do. It is very easy to make better health. the use of natural
vitamin supplements. Vitamins function in many metabolic reactions that
occur in foods consumed in the body, control of vitamins and energy
metabolism of our body. buy hydrocone
19-12-2013 om 22:30
geschreven door boxer
boxer
Preservation and promotion of health is achieved
through a combination of physical, mental and social well-being,
sometimes called "the triangle of health." Health is a positive concept
focuses on the social and personal resources, as well as physical
capacities. Buy Phentermine
19-12-2013 om 22:15
geschreven door boxer
buy lortab
I just got to this
amazing site not long ago. I was actually captured with the piece of
resources you have got here. Big thumbs up for making such wonderful blog
page!
This is like my fourth time stopping over your Blog. Normally, I
do not make comments on website, but I have to mention that this post really
pushed me to do so. Really great post .
19-12-2013 om 19:18
geschreven door buy valium roche
buy ritalin
Really i
appreciate the effort you made to share the knowledge.The topic here i found
was really effective to the topic which i was researching for a long time
I'm
still learning from you, but I'm trying to achieve my goals. I certainly
enjoy reading all that is posted on your blog.Keep the information coming. I
loved it!
Explore the evidence of early human behaviorfrom ancient footprints to stone tools and the earliest symbols and art along with similarities and differences in the behavior of other primate species.
From skeletons to teeth, early human fossils have been found of more than 6,000 individuals. Look into our digital 3-D collection and learn about fossil human species.
The layers that contain fossils and archeological clues can be dated by more than a dozen techniques that use the basic principles of physics, chemistry, and Earth sciences. Some techniques can even estimate the age of the ancient teeth and bones directly. Advances in dating have made human evolution very exciting
About six or seven million years ago there were no bipedal creatures resembling humanity. The earth was rich with diverse life forms, all subject to and shaped by natural forces and not the slightest signs of the intelligence and creativity that typifies humanity. Planet earth flourished without humanities' interfering nature as it had for billions of years.
A trend toward larger brain size, a primate characteristic, started with the australopiths. Food, its gathering and eating influences an animal's evolution (Reader, 1988). Primate species that eat higher-quality, more widely dispersed foods, generally have a larger brain. Brain enlargement probably arose in response to changes in foods (= habitat).
Two features that separate a humanoid form from an ape are bipedal locomotion and a larger brain volume. The largest known brain volume of the gorilla is 650 cc, and the smallest known in humans are 855 cubic centimeters.
Brain volumes of Australopithecus , the most primitive possible human ancestor identified, ranges from 435 to 650 cc (other say 413 to 530 cc), well within the gorilla and chimpanzee range. However, the part of the brain responsible for our greater mental dexterity than other primates, the cerebral cortex, is well developed in A. africanus (Dart, 1925) compared to other great apes.
By three million years ago, this creature already had expanded frontal lobes more human-like than that of any living primate (Falk, 2001).
Scientists include species within the Genus "Homo" and thus as ancestors of humanity on this basis. The "cerebral Rubicon" proposed is a cranial volume of 750 cc. Fossils called Java Man (850 cc) and Peking (915 to 1225 cc) are both included within the genus Homo. Homo habilis , the first species of our genus, had a brain volume of between 600 and 800 cubic centimeters (Wilson, 1992). Homo rudolfensis with a brain volume from about 775 cc to 900 cc, lived around the same time as H. habilis (Mayr, 2001).
Our upright method of walking evolved before significant brain enlargement occurred. In north-eastern Tanzania, at Laetoli is the oldest evidence of a bipedal (upright walking) hominid. These human-like footprints are found with guinea fowl and other bird prints, antelope and gazelle, giraffe, elephant, rhinoceros, pig, hyena, baboon, carnivores, hares, and three-toed horse (Leakey and Hay, 1979). Dated at 3.5 to 3.6 million years old, the prints are preserved in a volcanic ash bed (Reader, 1988). This is 1.6 million years before the oldest known tool use by early hominids! Some trails are 50 metres long and are entirely human, with a well developed arch to the foot! This creature's big toe appeared quite long, resembling the mobile toe of the chimpanzee, but the absence of any hand marks showed that the creature walked upright. Its stride suggests that the larger individual stood at 140 centimeters and the smaller at 120 centimeters. Some foot bones of such a creature have been found in South Africa (Sterkfontein caves). This creature's had an ankle bone adapted for upright walking and is almost identical to the ankle bone of a modern human.
The big toe angles away from the foot and the joints suggesting that it was highly mobile (Tobias, 1995). Scientists place these with Australopithecus (the "southern ape"), a hominid that lived in East and South Africa between 4.4 and 1.4 million years ago (" 1.4 to 4 lines ago ") (Avers, 1989). Sterkfontein would have been a steamy tropical jungle at the time the "Little Foot" entered Southern Africa.
Bipedalism is a major biological adaptation (Leakey, 1994). One question is whether the fossil record is complete enough and allows enough time for the evolution of Australopithecus sp. into Homo sp. Researchers can argue that some of the fossil features suggest that the Australopiths are not the direct ancestors of the genus Homo, but that they share a common ancestor. Some say there is too much biological discontinuity for the two creatures to be directly related. Others say that bipedalism requires important changes to the bones, muscle arrangement and limb movement.
Any difference, such as that of the inner ear bones (see details below), may be an evolutionary vestige of the animal's past. Researchers need more fossils, especially of Homo habilis , H. rudolfensis and the earlier species, such as Sahelanthropustchadensis, to resolve this problem.
Australopithecus was a bipedal ape with a small brain (450 cubic centimeters) (Washburn, 1978), and had massive, thickly enamelled molar teeth adapted to a diet of tough plant material. Its chest is funnel-shaped, with long arms and short legs as in apes. Bipedalismarose before the evolution of intelligence or large brain volumes associated with humans, a proposal that Haeckel presented in the 1860's (Reader, 1988). They may not have walked in the human manner but loped along similar to a chimpanzee. A naturally upright stance is further confirmed by foramen magnum and occipital condyles being set far forward and the neck muscles attach low down on the back.
There is also a prominent mastoid process , necessary for balance in upright walking. Bones of the inner ear of Australopithecus resemble those of the chimpanzees and gorillas more than humans, suggesting that either, these creatures were very early on the path of bipedalism or were not fully bipedal. Only in Homo erectus is the inner ear structure the same as in modern humans, showing that the creature definitely walked and ran exclusively on two feet (Horgan, 1994).
About 2.5 million years ago, several species of Australopithecus ranged in Africa from Ethiopia to South Africa.
Today scientists recognise numerous Australopithecine species (now called australopiths). A. robustus, is a heavily built creature, represented by specimens from southern and east Africa and existed 2.2 to 1.2 (a narrower 1.9 to 1.5 million years ago (Mya) by other estimates (Lemonick & Dorfman, 1999)) ( see timeline image ). A. africanus, the "gracile" species represented by the Taung specimen, lived 2.3 to 3 (or a wider 3.5 to 2) million years ago and A. afarensis, (e.g. "Lucy:) also a fine boned species, existing about 2.9 to 3.6 Mya (Lemonick & Dorfman, 1999) and is represented by fossils found in Tanzania and Ethiopia (A. aethiopicus). Sometimes they include another species A. boiseiin the classification (see skull ), of which fossils dated at 1.6 to 2.5 million years (1.4 to 2.3 mya by other estimates (Lemonick & Dorfman, 1999)) old have been found, but others say A. boisei is the same species as A. robustus (1.5 to 1.9 MYA). A. boisei was similar to robustus , but with even more massive (some molars being up to 2 cm across) face and cheek teeth (Mayr, 2001). The brain size is very similar to robustus , about 520 to 530 cc. A few experts consider boisei and robustus to be variants of the same species. A few people add other species, such as A. aethiopicus , (2.3 to 2.8 (or 2.6) Mya) from northern Kenya (usually included within A. boisei, as its ancestor). They have called another fossil group from Chad, the most western distribution of Australopiths, A. bahrelghazali, (3 to 3.5 Mya) (Gore, 1997) and another Ethiopian species A. garhi (2.5 MYA) (Lemonick & Dorfman, 1999). Most of the australopith males were between four and five feet tall and the females up to four feet tall.
Ardipithecus ramidus
The story of bipedalism and Australopithecus begins with the earliest known biped, Ardipithecus ramidus. This period covers the end of the Miocene epoch (5 to 23 Mya), the Pliocene epoch (1.8 to 5 Mya), the Pleistocene epoch (10,000 to 1.8 Mya) and up to today (Holocene epoch (zero to 10,000)). A few fossil fragments from Kenya are between 5.5 and 4.5 million years old (Bilsborough, 1992).
Since the chimp (Pan) and human lineages have split we have 4 recognised genera: Ardipithecus, Australopithecus, Paranthropus and Homo.
Remains of a creature in Ethiopia (Middle Awash/Aramis) , dated as living from 5.8 to 4.4 million years ago, called Ardipithecus ramidus(4.5 and 4.3 Myr ) ( Australopithecus ramidus (White et al., 1994)) and Ardipithecus kadabba(5.25.8 Myr), are probably ancestral to Australopithecus afarensis and subsequently the long line leading to humanity (and so the oldest hominid found to date).
Tim White of Berkeley, the anatomist working some individuals, says, "Ramidus is the first species this side of our common ancestor with chimpanzees."
Its similarities to chimpanzees includes the thinner tooth enamel and smaller molars of a fruit and vegetable eater, while its lower canines and upper premolars display hominid traits. All apes have large canines, while human canines are small. The brain volume of 'Ardi' was about the size of a modern chimpanzee.
A. ramidus was a forest dweller that does appear to have been bipedal due to the forward placement of the foramen magnum. It may have lived a partly arboreal existence, with ape-like grasping feet that facilitated tree climbing. Five to six million years ago, the Middle Awash region was deep rift valley with active volcanoes. The region would also have been showered by pulses of thick and hot volcanic ashes from these volcanoes. Ardipithecus lived in a high in elevation that was relatively cool, wet, and forested. Fossils of more than 60 mammal species have been found associated with the new hominid, including primitive elephants, rhinos, horses, rats, woodland forest antelope and monkeys (White et al., 1994). This species challenges the theory that the origins of bipedality evolved on the open savannah where greater distances needed to be quickly covered, or at least is the species that had already evolved a partially bipedal habit in a forested environment.
Hominids like Lucy (Australopithecus afarensis) were full bipeds, while Ardipithecus represents an evolutionary phase between our four-limbed palm-walking ape ancestors and the exclusive bipedality.
The small canine teeth in A. ramidus males, a characteristic, like bipedality, of all hominids, distinguishes them from great apes. Great ape canines are quite large, especially in males who use them in dominance displays, mating rivalries and occasionally defense from predators. Ardipithecus social-behavioral relationships must have evolved differently to those of other primates.
Ardipithecus represents an unspecialized ape evolving in the direction of Australopithecus, but with a mosaic of features, neither chimpanzee nor human. The limbs and hands resemble those of extinct apes and and not gorillas or chimpanzees.
Ardipithecus did not knuckle-walk as gorillas do, nor swing or hang from tree branches as chimps do; these representing later specialized traits of these two apes.
Australopithecus anamensis
Fossil remains of Australopithecus anamensis, have been found at Kanapoi and Allia Bay on the East African Rift System. This Australopithecine species is about 4.2 to 3.9 million years old. It was a bipedal ape and possible ancestor to A. afarensis(Gore, 1997). The dentition of A. anamensis is ape-like, with large canines, yet it was definitely a bipedal hominid, as seen from its tibia. A. anamansis was a small brained, biped (walked on two legs) with big teeth that fits into the one million-year gap between the earlier Ardipithecus and Australopithecus afarensis (which includes the famous fossil skeleton known as Lucy). A. anamansis fossils are anatomically intermediate between the earlier species Ardipithecus ramidus and the later species Australopithecus afarensis. Fossils of this species have been found in Ethiopia (Middle Awash area in the Afar desert of eastern Ethiopia) and Kenya. The Ethiopian fossils, at 4.1 milion years old represent a fossil continuity between Australopithecines and the earlier Ardipithecus species.
Hundreds of mammal fossils also were found in the region, indicating a woodland habitat with colobus monkeys, kudus, pigs, birds and rodents, and carnivores such as hyenas and big cats. This wooded habitat type persisted over a long period in this part of the Afar and was favored by early hominids between 4 and 6 million years ago.
A. afarensis
A. afarensis (3.9 to 3.0 Mya) (Eastern african species) precedes A. africanus (Southern African species) (3 to 2 Mya) by at least 500,000 years. Some scientists say that A. afarensis is probably the ancestor to every hominid fossil ever found. Fossil finds cover a period at least 1,000,000 years (3.8-2.8 Mya), suggesting a very long lived species. They became extinct about three million years ago (Avers, 1989). According to once scheme, this species is most likely the ancestor to two divergent lines:
Australopithecus afarensis (3.6 to 3.0 Mya) to A. africanus , to A. aethiopicus (2.6 Ma) to P. boisei (2.3 to 1.2 Ma) species and/or P. robustus lineage;
A route to humanity via A. garhi, then H. habilis , H. ergaster and H. erectus .
The more current proposal is that H. habilis is not ancestral to humanity. Another species, Homo rudolfensis, appeared in East Africa between 2.4 and 1.8 million years ago (Lemonick & Dorfman, 1999). H. rudolfensis appears to have migrated to eastern Africa from elsewhere. A new picture to the puzzle is emerging with the fossil finds, such as S. tchadensis, with its brow ridges and small canine teeth, which are characteristic of later hominids. H. rudolfensis may be ancestral to Homo ergaster ( erectus ). Homo erectus fossils have also been found in Algeria, Morocco and South Africa. Remains of Homo erectus and simple stone tools more than a million years old have been found in eastern Java, showing the wide range of this species (Bellwood, 1980). There is evidence that Homo erectus fossils found in Java are very old. One individual may be 1.8 million years old and another 1.66 million years old (Lewin, 1994). This species is also found in China and Indonesia.
These researchers believe that the human line (called Hominines or of the Genus Homo) diverged about 4 million years ago from the line that evolved into A. afarensis and Homo habilis. Until the discovery of Sahelanthropus tchadensis , there was no fossil record to support this (Parker, 1992) (Avers, 1989). Now, we have a possible ancestor to H. rudolfensis at nearly 7 million years ago. There is an increasing amount of circumstantial evidence in support of this view:
[1] Both Australopithecus and Homo sp. are bipedal apes as shown by the central position foramen magnum (the hole at the base of the skull for the spinal column), typical of an upright posture. This unusual feature leads to the two species being grouped together. Australopith members had much longer and curved toes (phalanges) and fingers. They also had a cranially orientated shoulder joint and other features of the arms, typical of tree climbers. As well as the foot, and foramen magnum, we see bipedalism in the shape of the pelvis and the angle between the thighbone and the knee (Leakey, 1994).
[2] A. robustus and A. africanus both had teeth different to the apes. Apes have teeth with pointed cusps as adaptations to a diet of soft fruit and other vegetation. The Australopiths have teeth flattened into grinding surfaces. This suggests a diet different to apes and consisting of tougher foods such as roots, nuts and hard fruits. Furthermore, the Australopiths did not have canines, as are found in other apes but not the Homo sp. Such dentition brings the classification of the Australopiths closer to the genus Homo. It is likely that the Australopiths ate mostly plant matter.
[3] In the genus Homo, the inner ear structure is identical to Homo sapiens sapiens (modern humans), while the Australopith's inner ear semicircular canals resemble those of apes.
[4] Australopith's pattern of growth and development was ape-like while early Homo erectus showed development midway between the ape and modern human (Leakey, 1994). As the brain capacity exceeds 770 cubic centimeters, serious constraints are encountered, as the pelvic opening becomes too small. A solution is found in human children being born earlier and with brains one third the adult size. Apes are born with brains half that of the adult size. This results in human babies being born more helpless.
[5] Australopith tooth development with age is ape-like, while H. erectus and the Neanderthals followed the human pattern.
[6] Australopiths have a conical shaped rib cage, similar to apes and tree climbers, not barrel shaped as in humans. Early Homo species must have been physically active, while this is not so with the Australopiths. A Homo erectus skeleton, called Turkana boy (KNM-WT 15000) (Brown et al. , 1985), was 1.64 to 1.68 metres tall and would have been powerfully muscled. Australopithecus would not have been able to breathe as we do when running. This places Homo as a runner and Australopithecus as a less active animal. To support this we find that Australopiths have a typical ape-like heavy build for their height, while early Homo species had the more agile build of humans.
[7] With the greater activity of Homo species as compared to the Australopiths, the blood vessel structure draining the brain of the Homo line is much more conducive to cooling than the Australopiths. An active animal has to remain cool and dissipate heat and its physiology has to adapt to this.
Fossil evidence such as a perfect knee joint shows that A. afarensis walked upright up to 3.6 million years ago. Bipedalism provides a competitive advantage as it is more energy efficient than quadrupedal locomotion. It also frees the hands for other activities (Chiras, 1994). The footbones of Australopiths are similar to those of the human foot, but have ape features that persisted with H. habilis . Its foot has long, curved toes, while the forelimbs are proportionally longer than in the genus Homo. A. afarensis has many cranial features reminiscent of our ape ancestry, such as a forward protruding (prognathic) face, a "U-shaped" (see AL 200-1) palate (with the cheek teeth parallel in rows to each other similar to an ape) and not the parabolic shape of a modern human's jaw teeth.
A female A. afarensis , "Lucy", (20 years old), with wisdom teeth , stood only 107 to 122 centimeters high. Its teeth showed a mixture of modern and primitive features. As with apes such as the chimpanzee, there is a distinct gap between the canines and incisors, the palate was shallow and the teeth rows were parallel rather than curved. An Australopith brain is barely larger than that of an ape of similar body size, such as a chimpanzee, yet it walked upright . Even at this early stage the brain structure differed from that of the apes, having a greater cerebral height, and more human-like frontal and temporal lobes. There is also evidence of the Broca's area, a part of the brain necessary for speech.
Lucy was less than one metre tall and probably weighed 27 to 30 kg, while some males topped 1.5 m and weighed up to 50 kg. This suggests that these creatures displayed sexual dimorphism. (Human males and females differ by an average of 13 cm.) Females appear to have been better at tree climbing while, the males were better at walking - a really odd differentiation. It's curved fingers were part of a creature that still climbed trees as a normal part of its life. Australopithecus ' erect bipedalism was probably not as developed, or identical to the perfection shown in Homo sapiens . Its upper torso possessed distinctly arboreal features and its leg's distinctly bipedal as expected from an ape newly on the bipedal road. Lucy's skeleton was different from earlier primates. Her knees were able to lock, her femur slanted inward and her large toe was in line with her other toes, allowing her to walk upright (see AL 129-1a/b) . Another feature that identifies bipedalism is the foramen magnum. This is the opening at the base of the skull through which the spinal cord passes. It is at the far back of the skull in quadrupeds (four-legged-walking) such as dogs, and is more centrally located in bipeds such as humans. In Australopithecus the foramen magnum is not quite central, so this creature may have walked with a stooped posture (Avers, 1989).
A. africanus
It is possibile that A. africanus and A afarensis ( see species time chart ) share a common ancestor and that A. africanus is the ancestor of H. habilis (Berger, et al , 1998) via A. garhi ( see above ) . This hypothesis arises from the incongruity that A. africanus, although a more recent species, had more ape-like longer arms and shorter legs that A. afarensis , from which it was supposed to have evolved. Further corroboration is found in the comparison of the thigh bone and arm lengths of Lucy (A. afarensis) and Homo habilis (and the discovery of A. garhi). The thigh bones are the same length, while the arms of H. habilis are longer. A. africanus also shares other features with Homo habilis . When compared to A. afarensis , it has smaller canine teeth, a larger brain and a shorter face. This raises the possibility that A. afarensis is not the ancestor of H. habilis , but more fossils are needed to confirm this. Lee Berger, the proponent of this idea even suggests that bipedalism may have evolved twice, one lineage in East Africa and another in southern Africa where a mix of habitats existed. Wear patterns on the teeth of A. africanus are similar to that of modern primates that eat leaves and fruit (Gore, 1997).
A. africanus represents thegracile fom of Australopith. All finds from South Africa are of around 2.4 to 3 MYA. Compared to robust Paranthropus species, their faces are more lightly built and dished-out, lacking a sagittal crest. Their cranial capacity of was about 450 cc.
A. garhi
Australopithecus garhi may represent an evolutionary link between the genera of Australopithecus and Homo (Asfaw et al 1999). It is probably descended from A. afarensis . It is associated with world's earliest stone tools found in the same general locality (Bouri, in the Middle Awash area in the Afar desert of Ethiopia) and dated, at 2.5 mya, and excavated antelope fossils with cutmarks made by stone tools and some opened by hammerstones (Sanders, 1999). However, the extremely large size of its teeth, especially the rear ones, and a primitive skull morphology make it unlikely to be a human ancestor according to other paleonologists.
A. garhi has larger postcanine dentition than A. afarensis. A. garhi lacks the dental, facial, and cranial features shared by A. aethiopicus, A. robustus, and A. boisei. Australopithecus garhi is distinguished from A. africanus and other early Homo species by its primitive frontal, facial, palatal, and subnasal morphology (Asfaw et al 1999).
Paranthropus species
After 2.5 mya, fossil finds show the Australopiths diverged in both east and south African populations into a line of gracile forms represented by A. africanus and a line robust forms as a different genus, Paranthropus (P. aethiopicus, P. robustus, P. boisei). The robust forms had larger cheek teeth (molars and premolars).
A. robustus , A. aethiopicus and A. boisei had a gorilla-like bony crest down the midline of the skull, serving to anchor the enormous jaw muscles needed to chew their fibrous diet (Wilson, 1992). This feature makes them look very ape-like. These robust forms were called "megadont " in that they had huge, broad cheek teeth with thick enamel (compare A. afarensis and P. boisei teeth from Asfaw, et al (1999)). Their incisor teeth relatively small. The rear teeth were designed for the stresses of heavy chewing and the associated skull morphology supports this observation. Their large zygomatic arches allowed the passage of large chewing muscles, while the large sagittal crest provided a large area to anchor these muscles to the skull. These are adaptations to chewing tough, fibrous foods.They also have very heavy brows making them look far from human. Anatomists group these three together as the genus Paranthropus and consider them an offshoot diverging from the ancestral line leading to humanity (Aiello & Dean, 1990). Scientists have not decided which Paranthropus species is closer to the ancestral human line. P. robustus, from South Africa (Swartkrans and Kromdraai) is the least 'human-looking' species, but has a human pattern of tooth development, while the tooth pattern of the other two species is distinctly ape-like. These species, P. boisei(Tanzania, Olduvai gorge) and P. aethiopicus, may represent evolutionary dead ends that separated from the Homo line at sometime.
P. robustus
P. robustus could be descended from the southern gracile A. africanus, in which case, the genus name Paranthropus would be invalid (see Paranthropus phylogeny). It lived from 2 million to 1.2 million years ago (others say 1.8 to 1.6 mya). This animal was around 1.1 - 1.3 metres tall, weighing between 40 and 80 kilograms and with a brain volume of 530 cc. It has a robust skull and grinding teeth. Its teeth indicate a diet of mostly coarse, tough food requiring a lot of chewing. It may have used tools. Fossils of this species are found in southern Africa.
P. aethiopicus
An example of this species is the skull KNM WT 17000, representing a distinct robust species, Paranthropus aethiopicus, that lived in eastern Africa 2.6 to 2.3 million years ago. It is possibly descended from A. afarensis, and ancestral to P. boisei, but could be an evolutionary dead end. This species is known from a fossil called the Black Skull and a few other specimens attributed to the same species. It has an unexplained mixture of primitive and advanced traits and a very small brain size at 410 cc. Parts of the skull are primitive and resemble A. afarensis. Other characteristics, like the massiveness of the face, jaws and the largest sagittal crest in any known hominid, place it close to A. boisei.
P. boisei
P. boisei (was Zinjanthropus boisei) persisted in east Africa from 2.1 until 1.2 mya. Fossils of P. boisei dated around 2 to 1.5 million years ago, are found with two species of Homo at Koobi Fora in Kenya. The exact identity of the two Homo species present is uncertain. Some classify them as Homo habilis and Homo rudolfensis (Groves, 1994). They have a larger brain and smaller teeth than Australopithecus and may be a single species (Gore, 1997). As anthropologists refine the study of these fossils, they are creating new species. Paranthropus walkeri is now seen as the link between australopiths and P. boisei (Groves, 1994) and P. crassidens from Swartkrans. This animal was around 1.2 - 1.4 metres tall, weighing between 40 and 80 kilograms and with a brain volume of 410 to 530 cc.
Kenyanthropus platyops
Another species of hominid, Kenyanthropus platyops (WT 40000) is dated at between 3.2 and 3.5 million years old. This places this creature in the same time period as A. afarensis . It has a flat face and small teeth. Its place in the human family tree has yet to be decided. Leakey et al (2001) see K. platyops as distinct from the australopithecines, sharing features with KNM-ER 1470 (Homo rudolfensis). It is possible that it is the ancestor of Homo, while the australopiths are a side-branch that are not ancestral to Homo. Tim White (2003) believes that the fossil skull is severely distorted and cannot be reliably identified. He says that it may be a Kenyan version of Australopithecus afarensis. (Image of K. platyops and H. rudolfensis from Lieberman (2001)).
The features that distinguish K. platyops from A. afarensis include primitive traits like small ear holes and advanced traits like a relatively flat face and small molars. K. platyops has morphological similarities with Homo rudolfensis ( KNM ER 1470) or Homo habilis, species that lived as many as 1 million years later. Similarities between these species might be explained in terms of convergent evolution, or by the possibility of a direct ancestral line between K. platyops and H. rudolfensis or H. habilis. Some scientists have placed H. rudolfensis specimens in the genus Kenyanthropus. Others ignore Kenyanthropus as a genus altogether, placing K. platyops specimens into the genus Australopithecus, saying that the species is not different enough to warrant its own genus.
Orrorin tugensis
Another bipedal hominid was discovered in 2000 by French anthropologists and named Orrorin tugenensis. At 6 million years old O. tugenensis ("Millennium man") is one of the oldest proposed human ancestors to date (Lemonick and Dorfman, 2001). It was estimate to be 1.2 metres tall and lived in a well-forested habitat. It has smaller molars and thicker enamel than the Australopithecine (now called australopiths) teeth, a characteristic of all later hominids. Richmond and Jungers of Stony Brook University published their findings in the March 21, 2008 issue of "Science" showing that the overall shape profile of the bone closely resembles early human fossils from about three to two million years ago; reflecting a bipedal animal. Biomechanically, O. tugenensis probably walked differently than modern humans.
The physical mechanism of upright walking remained the same from O. tugenensis to Australopithecus. Bipedalism is a key human adaptation and a defining featureof the hominin clade. O. tugenensis femur differs from those of apesand Homo and most strongly resembles those of Australopithecusand Paranthropus, indicating that O. tugenensis was bipedalbut is not more closely related to Homo than to Australopithecus.O. tugenensis shared distinctivehip biomechanics with australopiths, showing that this complexevolved early in human evolution and persisted for almost 4million years until modifications of the hip appeared in thelate Pliocene in early Homo (Richmond and Jungers, 2008).
About two million years ago, an environmental shift that occurred within East Africa, changing forests into grasslands, forced hominins to rely even more on walking as they looked for food. This added new selective pressures on bipedal walking leading to a dramatic change in hip structure. Longer-distance walking became necessary, giving humans the capabilities of today, with long striding legs and a very different kind of body shape compared to O. tugenensis or Australopithecus.
Another fascinating find is a male hominid skull from Chad, with a preliminary dating of 6 to 7 million years old, making it the oldest hominid fossil found, is nicknamed Toumai ( "hope of life" in the local Goran language). Sahelanthropustchadensis has a neck attachment to the skull from below, a feature of a bipedal animal, forcing a review of the origins of bipedalism. As no absolute date could be established, scientists had to use other fossils, such as pigs and elephants, found with the skull and that had been dated at other sites, to get an age estimate. This creature has a face similar to Homo habilis of 2 million years ago according to Dr. Daniel E. Lieberman, a Harvard palaeontologist. The Lucy fossil of 3.2 million years ago is very chimp-like by comparison! This suggests that the australopithecines (now called australopiths) my turn out to be a side branch outside the human ancestral line. Its face is "tall" with a massive brow ridge, while the mid-face is short (in the superoinferior dimension), being less prognathic than either Pan or Australopithecus (Brunet, et al. 2002). The teeth of the new fossils are taxonomically distinctive enough to assign the fossils to a new species and genus (Wood, 2002). Its brain volume was chimp-like at 320 to 380 cc. The site where S. tchadensis was found (TM 266) had a rich aquatic fauna of fish, crocodiles and amphibious mammals, alongside gallery forest and savannah animals such as primates, rodents, elephants, equids and bovids. This and sedimentological evidence, suggests that S. tchadensis lived close to a lake, and a sandy desert. The fauna suggests a biochronological age between 6 and 7 million years. This fossil could be close to the common ancestor of humans and chimpanzees.
The current evolutionary scenario emerging ( view timeline image ) is that, the original fossils identified as Homo habilis were as mix of species. However, the specimens described were too variable to belong to a single species. Larger-brained specimens have been renamed as H. rudolfensis (Mayr, 2001). Homo habilis is now considered to have evolved from the Australopithecines, but not to have evolved on to become Homo sapiens. It is even possible that tool use atributed to H. habilis, was really that of H. rudolfensis.
Sahelanthropus tchadensis may have been bipedal in its walking habits, but as bones from its legs and feet have not yet been found, this cannot yet be verified.
African Origins:
Australopithecus africanus lived in association with many African mammals. To the East of the Great Rift Valley is found the most ancient fossils of this species. This ape did not use fire or tools. Palaeontologists attribute tools found with fossil remains of Australopithecus sp . to Homo habilis or H. rudolfensis and the earliest tools known to date are about two million years old. Australopiths were about 1.5 metres tall (Wilson, 1992) and lived near rivers, streams and lakes in highland areas periodically inundated with volcanic ash from nearby eruptions. Fossil remains of crocodiles, flamingos, and Tilapia are commonly found with these hominid fossils. Environmental conditions fluctuated between wet and dry during this period. Evidence for this is from fossil pollen of vegetation that was present, the animal life found in each level and the wind-blown, or water-borne materials.
Yves Coppens, a palaeontologist, did some important detective work on the site of our origins. His proposal is founded strongly upon geology, molecular biochemistry (molecular biology), palaeontology and ecology. His is the type of reasoning that emerges once we accept that we are evolved animals. Molecular biologists have established that humanity is closely related to the chimpanzee and that lines such as the Orangutan are very distantly related to humanity. With palaeontologists, they came to a "prehistoric compromise", agreeing that the lineage leading to humanity separated from the chimpanzee lineage seven and a half million years ago.
An objection was that if Pan , the chimpanzee was our ancestor, why were no such fossils found with Australopithecus in eastern Africa. Hominid fossils (family Hominidae) as old as seven or eight million years have been found in Ethiopia, Kenya and Tanzania, but no chimpanzee or gorilla ancestors (family Panidae) occur in the same area. Molecular biology, biochemistry and cytogenetics all show that humans and chimpanzees evolved from a common ancestor. A solution to this puzzle came from vertebrate distribution maps and geological history. Africa's Rift Valley runs from north to south. Vegetation and climate differ dramatically on each side of this rift. To the west are woods and forest where chimpanzees are found and to the east are grasslands where only hominid fossils are found.
The hominid and chimpanzee ancestor lived in a homogeneous biogeographical province, a massive lush forest in central Africa. Roughly twenty million years ago, the Rift valley started to divide equatorial Africa due to east-west stresses in the tectonic plates (Maslin, 1994). According to geophysicists, this tectonic activity created the 2000 metre high Kenyan and Ethiopian domes. Rainfall patterns began to alter and rainforest, woodlands, shrub and grasslands developed in close association. As the rift subsided into a large valley, a process evolving over ten million years, the Eden of this ancestral species split into two. This new geological structure was a natural barrier that altered the weather patterns, with the west remaining humid and moist. Eastern Africa became organised into a seasonal monsoon and according to palaeoclimatologists, progressively drier. Within the unchanged western forests and woodlands the chimpanzee evolved from this common ancestor. On the east the change in climate subjected this ancestor to different selective pressures and a more drastic change in this creature's habitat and niche. Forest gave way to open savannah. Palaeontologists note a change of species to what they call Ethiopian fauna, eight to ten million years ago. This was accompanied by a decline in the number of ape species from more than 20.
Along the African Rift System, which runs from the Red Sea in the north to Lake Malawi in the South a diverse habitat evolved, with ponds, small streams, woodlands, savannah plains, and rocky valleys. Along this valley and extending down to Sterkfontein and Taung in Southern Africa, are the many fossil sites for the Australopiths and early Homo species. Natural selection driven by climatic and vegetation changes favoured a less arboreal way of life, suited to the bipedal savannah ape, Australopithecus . As food was more dispersed bipedal apes could forage more successfully . As with much species formation, the genetic divergence of our ancestor from the line leading to the chimpanzee, arose through geographic isolation caused by the formation of the Great Rift Valley. The process was also driven by a shift toward prolonged and seasonally more arid conditions, opening up grasslands in place of forests after 2.8 million years ago (deMenocal, 1995). The chimpanzee ancestor remained in the available forest habitat and became the species if chimps we find today. Another lineage from this common ancestor adapted to the more open grasslands, experiencing completely different selective pressures, evolving to occupy a comletely different habitat - open graslands.
As an example of other evolutionary trends that follow through from Australopiths, we find that there is strong sexual dimorphism in the Australopiths, similar to what exists in gorillas and orangutans today. There is good evidence of large sex differences in A. afarensis (Leutenegger, 1995). This is clearly seen in the jaws, skulls and the thigh bone. A pattern of sexual dimorphism continues through to Homo habilis and Homo erectus , but is reduced in archaic Homo sapiens and Neanderthals. Its reduction in modern humans is due to females getting larger and not males getting smaller. This difference seems to vary with environmental and nutritional conditions. Dutch women of 100 years ago were about 10 centimeters shorter than males. Less that 2 percent of males reached 180 cm then. Today the average Dutchman is 183 cm tall and 14 cm taller than the average woman (Usher, 1996).
We do not have the fossil remains of our ancestors of between 5.5 and 10 million years ago. There is no fossil record for the African apes (Gorilla and Chimpanzee) due to their living in moist tropical forests where decomposition is rapid, preventing fossilization. It follows that there is a limit to how far back in time hominid fossils will be found, as we originated in a similar habitat. Molecular comparison of living primates suggests that the African apes, the Chimpanzee and Gorilla, split from the hominid line between 6 and 8 million years ago (Mya) (Aiella & Dean, 1990).
Robust Australopiths were vegetarians, with heavy, specialised teeth. Such animals would not have much use for complex tools. This diet doomed them to extinction as the evolutionary pressures of this route forced greater and greater morphological specialisation. We see a trend in the increasingly specialised characteristics of the line, leading to a branch yielding A. africanus about 2.5 million years ago, then to A. robustus and then A. boisei , without selecting for a larger brain and finally becoming extinct. Their facial architecture becomes increasingly bulky through time, selected for greater chewing forces to handle their herbivorous diet. Their adaptive solution to selective pressures was morphological instead of behavioural. Herbivores typically lack the intellectual challenges faced by predators and do not need a cooperative culture.
Tools?
Australopiths did not shape stone tools, but if they used wooden tools such as a stick, no fossil evidence would remain. Our closest living relative, the chimpanzee, uses tools spontaneously, both in the wild and in nature. Wooden tools will not fossilize and chimpanzees do not shape stone tools for use. Chimpanzees use many different tools, such as "dipping" for ants or termites using a stick, using a leaf as a sponge and using a stick as a weapon. Weapon use, shown mostly by males, involved sticks and stones that they flailed, clubbed and threw at other chimpanzees, humans and baboons. In one experiment, a chimpanzee attacked a stuffed leopard, using a stick as a club. It hit the leopard with enough force to break a live leopard's back! (Jolly, 1972). Researchers list 19 habitual patterns of tool-use (McGrew, 1993). More than 34 populations of free-ranging, wild chimpanzees have different repertoires of tool use. These chimpanzees learn by imitation, so that even in the wild there are population differences in tool-using traditions (Gibson, 1993). Technology transfer even occurred, where a chimpanzee moved from one community to another and introduced the use of twigs to catch carpenter ants (Miller, 1995).
Chimpanzees of the Tai Forest are impressive. A mother was observed showing her five-year old child the necessary spatial orientation needed to hammer open a nut ( Coula edulis and Panda oleosa and three less common species) with a tool. The juvenile then attempted, copying the mother. To open a nut, a mother chimpanzee carries some nuts to a root, which she uses as an anvil. She places a nut in a depression on the root and hits it with a hammer tool - a wooden club or stone. Wooden sticks are prepared in various ways before use, such as shortening the stick to a suitable length. The harder the nut, the heavier and harder the hammer. Panda oleosa , is the hardest nut, so Chimps transport tools in foraging for this nut (Boesch, 1993). Rare stone tools are more often transported. Chimps remember up to five different tool locations and go to the closest, usually from a distance of at least 100m. Here we see how a specific diet, based on a single tree species, led to unique behavioural adaptations in the chimpanzee.
Mother chimpanzees also gave their child a good hammer. She would leave nuts near the anvil or put the nut on the anvil and leave the tool nearby, so providing an easy incentive for the young animal to try to crack the nut. Active teaching even took place. Panda oleosa nuts are the hardest and most difficult to open. They have three kernels independently embedded in a hard wooden shell. A partly opened nut had to be carefully and skilfully repositioned to get each kernel without smashing them. A young male succeeded in removing one kernel but then incorrectly placed the nut for the attempt to get at the second kernel. The mother quickly picked up the nut, cleaned the anvil, and replaced the nut correctly. The youth then succeeded in opening the second kernel.
In another episode, a young female was having trouble opening nuts with an irregularly shaped tool. She changed her position about 14 times and her grip on the tool about 40 times. She also tried moving the nut to a new position and rotating the nut to hit it on a different position. Having exhausted all the imaginable variables over an eight minute period, her mother, who had been resting, joined her. With the daughter sitting in front, she slowly rotated the hammer to the best grip, taking about one minute to do this and then opened 10 nuts. The daughter ate all the contents and then took the hammer. She then opened four nuts in 15 minutes, so still had difficulties, but always held the hammer in the position shown to her. At each difficulty she whimpered for attention and threw a tantrum after unsuccessfully trying to open the fifth nut for three minutes.
The nut-cracking techniques of this population of chimpanzees take 10 years of practice to master, so such tool-use could only develop where the nutritional rewards are high to compensate for the long period of learning required. This is a long period of trial for a consistent or perceived reward. Nuts can be cracked with the chimpanzees own teeth, but tool use permits individual chimpanzees to process more nuts than they can consume. Mother chimpanzees that master this nut cracking technique, provide the excess, up to 1000 calories per day in nuts, to their young until they reach about eight years of age. This gives the young extra and valuable nutrition and the leisure time to practice and develop their own tool-using skills.
In captivity, chimpanzees also use tools. When researchers hung food in their enclosure, they acted as anthropoid engineers, stacking up crates and using a pole to reach suspended food. Chimps solved the problem without help, using available materials. In this way, chimpanzees are the only nonhuman species in nature to associate and use different tools to solve problems (Mc Grew, 1993). A chimpanzee in the wild became the dominant male through his use of empty kerosene cans that he banged together to intimidate larger males (Miller, 1995).
Human capabilities far outreach this behaviour, but the behaviour in a nonhuman provides credence to the possibility of crude tool use in our most ancient ancestor. This social and technological complex in the chimpanzee provides a window to our most ancient beginnings. Jane Goodall noted a wide complex of chimpanzee behaviour during 35 years of study. This included meat eating, toolmaking, deliberate planning, warfare, cannibalism, the formation of coalitions, consortship, adoption of non related chimps, exploitation of social situations, the use of medicinal plants, and technology transfer (Miller, 1995).
From an evolutionary point of view there is great advantage in obtaining an excess of a highly valuable food resource. Nut cracking provided a net benefit of 3800 calories per day during the nut seasons (Boesch, 1993). A population of chimpanzees could develop in two specific directions. One could rely on its teeth to crack nuts and another on its ability to learn a transmitted skill. Natural selection would act on these two traits - "favouring" tooth jaw muscle development in the one and mental dexterity in the other. Nut cracking behaviour using tools was first reported by American missionaries working in West Africa (only found in Liberia and Ivory coast) in 1843, so there is little chance that humans somehow transmitted this behaviour to the chimpanzees (Hewes, 1993).
Another significant component to this chimpanzee nut cracking it that they would leave stone tools that worked well as nut cracking hammers and anvils at the tree base. They would revisit and use the same tool when the nuts in that oil-palm tree ripened. These chimps had a "mental map" of both where the oil-palm trees and their stone tools were. When another chimp has taken the tool, the returning chimp would search for its missing tool in the vicinity where it was last found. A chimp would take the tool with him to the next tree, so would have to remember at which tree the tool was last discarded (Potts, 1996). With this behaviour of bringing rocks directly to the food source, natural selection comes into play on various features:
The tool replaces teeth as a food processing organ, so selection will favour dexterity in tool use.
The chimp that better remembers when trees ripen and where the trees and tools are will fine life "easier"; a selective advantage.
Australopithecus's jaws have reduced canines and incisors and large molars and premolars. Their teeth show heavy wear. Its a strong projecting face and long zygomatic arch concentrates chewing power over the cheek teeth. With such a specialised chewing apparatus and large jaw muscles, they probably ate tough seeds or nuts. The tooth wear of the g
1.- About 90% of primate fossils consist of teeth, often just a scrap of one or two molars. Human and ape molars, thats gorillas, chimps, gibbons, o-rangs, and us, have a distinctive series of five cusps referred to as Y-5. A single molar, like the one shown left (by paleo-wildlife artist Carl Buell,) tells a paleoanthropologist theyre dealing with either a great ape or a hominid. It looks like there's plenty of fossil teeth for Ardi, I infer she passes this hurdle with ease
2.-.
Another important diagnostic trait is the spine, Human spines have an elongated "S" appearance when viewed from the side, that gentle double curve is unique in all the animal kingdom. It is found only in bipedal primates, i.e., hominids. All a scientist needs is two or three vertebrae from various sections to determine if the creature was bipedal.
3.-
In addition, the skulls of apes and hominids differ markedly underneath, where the spine inserts in the braincase. In humans and our close bipedal relatives, that hole, called the foramen magnum, is directly below the cranium so that the spine fits in at a 90 degree angle. In apes, its further back. The two skulls to the left, ( PZ Myers, )show that opening and its position in a human (top) and a gorilla (bottom). Looking at the fossil reconstructions, it appears to me that Ardi's spine passes bipedal muster, and her foramen mag is located either directly under her skull like ours, or just slightly farther back.
4.- Bipeds have distinctive pelvises, femurs, tibias, and feet. Shown right, from left to right, is the pelvis and femur/knee of a chimp, Lucy, and a modern human. Note how the chimp pelvis is long and flat, and the femur goes straight down. Whereas Lucy's pelvis is more compact and the femur fits in -- what anthropologist call articulates -- in such a way that the thigh juts in toward the opposite knee. The tibia, or shinbone, articulates with the femur at that knee joint at a slight angle compared to the chimp's as well, meaning the legs are 'tucked in; underneath the bipedal Lucy and modern humans. And while we like to think of our tool making hands as a defining human trait, it is our feet that have been radically reworked by evolution, from prehensile grasping digits with a thumb-like opposing piggy-that-went-to-market, to a robust, arched platform to stride heel-to-big-toe across the hot, flat, Pleistocene plains. In the video on the Science page, it sounds like Ardi's feet are flat, or flatter anyway. But she walked on those flat soles, not on curled up knuckles like a gorilla or a chimp. And her big toe is intermediate, it's not completely opposing, like an ape, but it sticks out at a 45 degree angle, so it's not in line with the rest of the toes, like ours or Lucy's
5.- .
Chimp Ardipithecus Afarensis (Lucy)
So here we see the skeleton of a chimp, a front and side view of Ardi, and Lucy aka Astralopithecus afarensis, roughly to scale (Ardi is just over one-meter or a little over three feet in height). And low and behold, Ardi has the double curved spine of a hominid, long ape-like arms, while pelvis, legs, and feet are almost perfectly in between. Based on all that, Ardi could walk upright almost as well as Lucy, using her arms to carry back goodies from the grassland to her forest enclave. And with those long arms, fingers, and toes, including the partially opposing big toe, she could probably climb almost as well as an adult chimp or juvenile gorilla. That's about as good a transitional fossil between a more ape-like, knuckle-walking ancestor and a bipedal hominid like Lucy as you could ask for.
06-10-2009 om 10:15
geschreven door tsjok45
04-10-2009
How will creationist handle this ?
1.-How will creationist handle this ? The same way they always do. The Creationists will find a way to make excuses for this find. Theyll declare it a hoax, or decide its entirely human, or entirely ape; -Definitely an Ape Its the same kind as a chimp. So was Lucy. It looks like nothing more than a very old gorilla. Perhaps a type that when extinct centuries ago. Do these scientists ever think of that?
-Definitely a Human just like Homo erectus and Homo neanderthalensis were of the same kind as humans. or theyll simply ignore it. No amount of evidence will convince these people
2-the "evolutionist Darwinistic scientists" themselves declare : " humans do not descent from chimps or other "apes" " ( See O'brian ) : Ignore the fact that the primary papers themselves dont make such claims and instead focus on the sensational press headlines. 3.- Modern human skeletons from the same site, so it lived at same time and was not an ancestor (Java man argument) 4.Never mind the continuity found in all hominimen fossils . Thats all a big cosmic joke (to temp our faith, I suppose).
( in other words ) creationists will answer by a combination of lies, distortion and ignorance! Theres no reason to assume they will change their usual tactics for this one!
Is ID de wetenschappelijke (sic) uitleg voor creationiisten 150jaar Darwinisme... De evolutietheorie gewikt en gewogen David sorensen en revolutietheorie lalala DE VERRIJZENIS VAN DE JONGE VERDRONKEN KOE » Reageer (21) ID TOVERDOOS OVER SLECHTE ID-EETJES ,RESISTENTIE & TOVERDOCTORS c) OEC d) YEC
Een oeroud spoor Denisova : ZUID SIBERIË DENISOVA - mens
DMANISI AAP OF MENS DE OUDSTE ? » Reageer (1)Nakalipithecus nakayamai (<) Alweer eentje ? ..... De tand des tijds / Heidelberg-mens DE HEUPEN VAN EVA CASABLANCA MAN /erectus Hobbit is aparte soort ? » Reageer (2) NOG EEN BENDE BIJTERS POLONAISE met neanderthaler TANDEN UIT DE QESEM GROT Neanderthaler genoom Mitochondriale Genenkaart van Neanderthaler Xuchang mens
b)Biologie 1.-(EVOLUTIE ) NAS / IM Document 2008 (2) NAS / IM Document 2008 ( 1)
Cambrium & precambrium AVALON GABONESE chips
Evolutie in actie Opmerkelijk snelle adaptaties bij kroatische ruine-hagedissen:
EEN PLUIM VOOR CHINA Geef eens een pootje Pluimgewicht KLEIN DUIMPJE & DE REUS &VEREN Opnieuw gevederde Maniraptor Similicaudipteryx. Aerosteon riocoloradensis
Krokodillen
PAKASUCHUS KAPILIMAI Prestosuchus chiniquensis Terug naar zee viseter
Eritherium azzouzorum LJOEBA Darwinius masillae VLIEGENDE KATTEN ? Vleermuizen ALWEER EEN BELANGRIJK STUK UIT DE LEGPUZZEL Zee-zoogdieren ; Van de wal in de visgronden Indohyus / walvisevolutie Zeehonden-evolutie
*Blauwe en groene ( =nederlandse)teksten zijn meestal aanklikbare links *Engelse Wikipedia teksten verwijzen in de linkerkolom naar verschillende niet-engelse versies van het wikiartikel *Blauwe teksten tusssen "" , zijn voornamelijk ( gedeeltelijke)citaten afkomstig van mensen met andere meningen
Evolutie / Charles Darwin aan de basis: SELECTIE NATUURLIJK DE LEVENSBOOM UITSTERVEN DEEP TIME Biogeografie SEKSUELE SELECTIE Al jaren lang samen CO-EVOLUTIE De economie van de natuur Geleidelijke veranderingen
Unintelligent design:
IP Theorie Ons gebit , De appendix, Geboorte, De weg van het zaad , Nervus Laryngeus Recurrens, DNA , 75 mistakes , Het boek
Unintelligent design (1): Rechtop lopen Tomaso over het oog , oogspieren , Unintelligent Design (5): ZWEETVOETEN Unintelligent design (6): Vitamine C Unintelligent design (8): Kuitspieren
 Unintelligent design (1): Rechtop lopen
Unintelligent design (1): Rechtop lopen





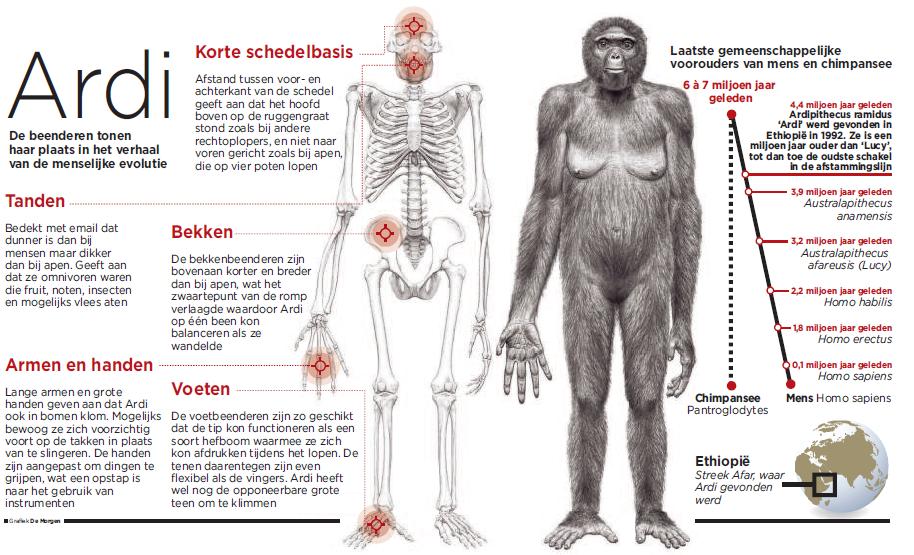
















 Fossil remains of
Fossil remains of  A. robustus , A. aethiopicus and A. boisei had a gorilla-like
A. robustus , A. aethiopicus and A. boisei had a gorilla-like  Another bipedal hominid was discovered in 2000 by French anthropologists and named Orrorin tugenensis. At 6 million years old O. tugenensis ("Millennium man") is one of the oldest proposed human ancestors to date (Lemonick and Dorfman, 2001). It was estimate to be 1.2 metres tall and lived in a well-forested habitat. It has smaller molars and thicker enamel than the Australopithecine (now called australopiths) teeth, a characteristic of all later hominids. Richmond and Jungers of Stony Brook University published their findings in the March 21, 2008 issue of "Science" showing that the overall shape profile of the bone closely resembles early human fossils from about three to two million years ago; reflecting a bipedal animal. Biomechanically,
Another bipedal hominid was discovered in 2000 by French anthropologists and named Orrorin tugenensis. At 6 million years old O. tugenensis ("Millennium man") is one of the oldest proposed human ancestors to date (Lemonick and Dorfman, 2001). It was estimate to be 1.2 metres tall and lived in a well-forested habitat. It has smaller molars and thicker enamel than the Australopithecine (now called australopiths) teeth, a characteristic of all later hominids. Richmond and Jungers of Stony Brook University published their findings in the March 21, 2008 issue of "Science" showing that the overall shape profile of the bone closely resembles early human fossils from about three to two million years ago; reflecting a bipedal animal. Biomechanically,  Another fascinating find is a male hominid skull from Chad, with a preliminary dating of 6 to 7 million years old, making it the oldest hominid fossil found, is nicknamed Toumai ( "hope of life" in the local Goran language).
Another fascinating find is a male hominid skull from Chad, with a preliminary dating of 6 to 7 million years old, making it the oldest hominid fossil found, is nicknamed Toumai ( "hope of life" in the local Goran language).  A single molar, like the one shown left (by paleo-wildlife artist Carl Buell,) tells a paleoanthropologist theyre dealing with either a great ape or a hominid. It looks like there's plenty of fossil teeth for Ardi, I infer she passes this hurdle with ease
A single molar, like the one shown left (by paleo-wildlife artist Carl Buell,) tells a paleoanthropologist theyre dealing with either a great ape or a hominid. It looks like there's plenty of fossil teeth for Ardi, I infer she passes this hurdle with ease

 Bipeds have distinctive pelvises, femurs, tibias, and feet. Shown right, from left to right, is the pelvis and femur/knee of a chimp, Lucy, and a modern human. Note how the chimp pelvis is long and flat, and the femur goes straight down. Whereas Lucy's pelvis is more compact and the femur fits in -- what anthropologist call articulates -- in such a way that the thigh juts in toward the opposite knee. The tibia, or shinbone, articulates with the femur at that knee joint at a slight angle compared to the chimp's as well, meaning the legs are 'tucked in; underneath the bipedal Lucy and modern humans. And while we like to think of our tool making hands as a defining human trait, it is our feet that have been radically reworked by evolution, from prehensile grasping digits with a thumb-like opposing piggy-that-went-to-market, to a robust, arched platform to stride heel-to-big-toe across the hot, flat, Pleistocene plains. In the video on the Science page, it sounds like Ardi's feet are flat, or flatter anyway. But she walked on those flat soles, not on curled up knuckles like a gorilla or a chimp. And her big toe is intermediate, it's not completely opposing, like an ape, but it sticks out at a 45 degree angle, so it's not in line with the rest of the toes, like ours or Lucy's
Bipeds have distinctive pelvises, femurs, tibias, and feet. Shown right, from left to right, is the pelvis and femur/knee of a chimp, Lucy, and a modern human. Note how the chimp pelvis is long and flat, and the femur goes straight down. Whereas Lucy's pelvis is more compact and the femur fits in -- what anthropologist call articulates -- in such a way that the thigh juts in toward the opposite knee. The tibia, or shinbone, articulates with the femur at that knee joint at a slight angle compared to the chimp's as well, meaning the legs are 'tucked in; underneath the bipedal Lucy and modern humans. And while we like to think of our tool making hands as a defining human trait, it is our feet that have been radically reworked by evolution, from prehensile grasping digits with a thumb-like opposing piggy-that-went-to-market, to a robust, arched platform to stride heel-to-big-toe across the hot, flat, Pleistocene plains. In the video on the Science page, it sounds like Ardi's feet are flat, or flatter anyway. But she walked on those flat soles, not on curled up knuckles like a gorilla or a chimp. And her big toe is intermediate, it's not completely opposing, like an ape, but it sticks out at a 45 degree angle, so it's not in line with the rest of the toes, like ours or Lucy's








 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
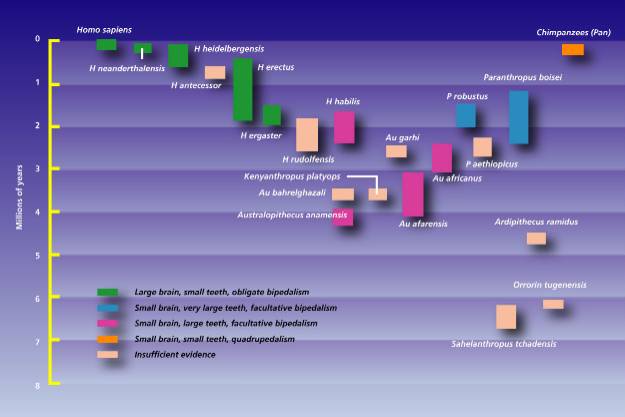{kind=link}
{kind=link}