Dit blog is een onderdeel van EVODISKU WAT IS DE BEDOELING EN WAAR STAAT DIT BLOG VOOR
****Wie meent dat alles inmiddels wel over de evolutietheorie gezegd is en dat de discussie gesloten kan worden, ziet over het hoofd dat de wetenschap niet stil staat.
***Wie meent dat inmiddels het creationisme definitief het pleit heeft verloren en dat de discussie gesloten kan worden , ziet over het hoofd dat het "creationisme" is geevolueerd ( en zal evolueren ) in nieuwere mimicrytische vormen( meme-complexen ) zoals bijvoorbeeld het ID(C)
***Dit blog is speciaal opgezet om de aktualiteit binnen het evolutie-creationisme debat te volgen en van kommentaren te voorzien ... waartoe de lezers zijn uitgenodigd bij te dragen ...
Let echter wél op het volgende :
"Je bent een rund als je hier met religie stunt " ....
21-12-2007
Van de wal in de visgronden
INDOHYUS
WAARSCHUWING Zoals steeds stonden de ( nederlandstalige ) nieuws-media weer vol met gejubbel , conclusion jumping en verschrikkelijke pop-wetenschappelarij , met ronkende titels en verhaaltjes over sensationele nieuwe "ontdekkingen " ...Het zou gaan om een langgezochte "missing link " die "uiteindelijk " is gevonden tussen walvissen en landdieren _____en nog het meest van al over een zeer oppervlakkige lezing van het artikel van Thewissen zelf
Ik was er natuurlijk ook ingetrapt , toen het bericht voor de eerste keer verscheen op het net.....:oops: ( gelukkig zette Noorderlicht/ me uiteindelijk op het goede spoor ...en vandaag verscheen ook het lezenswaardige doordachte en goed nagecheckte artikel van de NRC )
( artistic view/reconstructie ) olieverf schildering ( Carl Buell ) De zware botten en chemische samenstelling van de tanden laten zien dat Indohyus veel tijd onder water doorbracht. Hier duikt het hoefdiertje in een stroom zoals de Afrikaanse kantjil tegenwoordig doet als er gevaar dreigt.
(Het afbeelden van luchtbellen is een onnodig fantasietje (de afgebeelde vacht is dat evenzeer )... we weten NIET of het dier het uitademen ( =het lozen van uitademhalingsgassen ) onderwater , al dan niet kon uitstellen ....Dit is op zijn minst een ongeoorloofd detail in deze "artistieke" reconstructie ) 'Indohyus' is geen walvis. Zou je ook niet zeggen, want Indohyus heeft nog het meeste weg van eenkantjilof dwerghert.
Toch is ie familie. Sterker nog, volgens Thewissen is Indohyus, het met de walvissen nauw verwant beest. ( zei hieronder in (A) : Thewissen zelf over deze verwantschappen ) Echter niet de voorouder, want Indohyus is 'slechts' veertig miljoen jaar oud en de walvissenfamilie ontstond vijftig miljoen jaar geleden al. (1) Toch zal de echte voorvader van de walvissen wel enigszins op Indohyus geleken hebben.
De Indohyus,( van de Raoellidae - groep )was een klein vierpotig herbivoor land-zoogdier dat vrij regelmatig in (en onder) het water vertoefde De auteurs van deze nieuwe studie stellen voor dat de vroegste voorouders van de walvissen op dezelfde manier leefden ....(1) Gradueel veranderde de walvisstamlijn in carnivore , meer aan het water-millieu aangepaste afstammelingen ; zoals bijvoorbeeld ;, Ambulocetus, die men zich het best kan voorstellen ales een grote behaarde alligator ( zie onderaan dit artikel voor het holotype-fossiel) ...
Hans Thewissen over deze vondst en over walvissenevolutie ---> this video
Amerikaanse onderzoekers hebben een mogelijke voorouder van een neef van de walvissen gevonden die gedeeltelijk in het water is gaan leven ( bijvoorbeeld om aan roofdieren te ontsnappen ) en aanvankelijk leefde van lanplanten
Wetenschappers van de Northeastern Ohio Universities College of Medicine hebben fossiele resten gevonden in de streek Kashmir in India Het zijn de overblijfselen van een hertachtig diertje ( ongeveer zo groot als een vos /of zo groot als een " wasbeer (2 ) dat indohyus is gedoopt ... Het onderzoek ( *1 ) vond aanwijzingen dat dit de meest directe (tot nu toe bekende ) verwant is van de walvisachtigen :
Het was al geweten dat walvissen afstammen van een landdier De Indohyus lijkt dan wel in niets op de walvissen zoals wij ze vandaag kennen, toch zijn er overeenkomsten.
De Indohyus leefde 48 miljoen jaar geleden en was een vertegenwoordiger van de Evenhoevigezoogdieren . Indohyus behoort tot de oude tak van zoogdieren die twee of vier tenen hadden aan elke voet. Onder de afstammelingen van die tak rekent men vandaag kamelen, varkens en nijlpaarden. ( orde ; artiodactyla --> zie hieronder appendix )
a, b, Oblique lateral view of skull RR 208 (a) and ventral view of skull RR 207 (b).
ch, Posterior views of humerus (RR 149, c) and femur (RR 101, d), plantar views of metacarpal (RR 138, e) and proximal manual phalanx (RR 19, f), dorsal view of astragalus (RR 224, g), and posterior view of metatarsal (RR 139, h). il, Histological mid-shaft sections for humerus of the pakicetid Ichthyolestes (H-GSP 96227, i), humerus of Indohyus (RR 157, j), femur of Indohyus (RR 42, k) and femur of the artiodactyl Cainotherium (IVAU unnum, l). Both scale bars are 1 cm; the scale bar near d goes with ah, and that near l goes with il.
The inner ear looks very whale-like
.....Indohyus shares with cetaceans several synapomorphies that are not present in other artiodactyls. Most significantly, Indohyus has a thickened medial lip of its auditory bulla, the involucrum (Figs 1 and 3), a feature previously thought to be present exclusively in cetaceans. Involucrum size varies among cetaceans, but the relative thickness of medial and lateral walls of the tympanic of Indohyus is clearly within the range of that of cetaceans and is well outside the range of other cetartiodactyls (Fig. 3). Other significant derived similarities between Indohyus and cetaceans include the anteroposterior arrangement of incisors in the jaw, and the high crowns in the posterior premolars.
fig 3
In cetaceans (open squares), the medial tympanic wall is inflated and called the involucrum, and the lateral tympanic wall is thinned and called the tympanic plate. In artiodactyls (open triangles), the medial and lateral tympanic walls are more similar in thickness, causing values on the y axis to be closer to 1. See Supplementary Information for further details.
Waterdieren
De structuur van ( vooral het middenoor ) is een belangrijke determinatiesleutel voor extante en extincte walvisschedels en uiteraard ook voor de studie van de evolutie van de walvissen
Het fossiel is al 15 jaar geleden ontdekt en geborgen , maar toen onlangs per ongeluk tijdens een onderhoudsbeurt van het fossiel ,een stuk oor afbrak , kwam het middenoor prominent bloot te liggen ...Thewissen merkte de gelijkenis met de oorstructuur van de oerwalvisssen gelijk op ....
Het is onder andere de oorstructuur (= the medial tympanic wall is inflated and called the involucrum, and the lateral tympanic wall is thinned and called the tympanic plate).en de tandmorfologie die Indohyus tot familie van de walvissen maakt. ( zie ook ) evolutie walvis oor <
*In de schedel van deze Indohyus zit een verdikking( involucrum ) rond het middenoor, net zoals bij voorhistorische walvissen. (3-)
*Tegelijkertijd heeft het dier zware botten, een kenmerk dat ook bij nijlpaarden voorkomt en dat moet vermijden dat het dier moeilijk moest manoeuvreren bij langdurig verblijf in het water : dikke botten zorgen immers ook voor een beter evenwicht in ondiepe wateren, zoals bij andere waterdieren. Ze zorgen tevens voor ballast zodat ze beter onder water kunnen duiken en onderwater-zwemmen ...
Er zijn in de skelet overblijfselen ook isotopen-verhoudingen gevonden die duiden op waterdieren ... M.a.w. de botten zijn osteosclerotisch (grote hoeveelheid compact bot waardoor de botten zwaarder zijn en het mogelijk is om beter in water te waden)
Results are shown as means s.d. for the sample population. Filled triangle, Indohyus; open triangle, Khirtharia; open squares, terrestrial mammals; filled squares, brackish-water anthracobunids; filled circles, freshwater/brackish-water archaeocetes. See Supplementary Information for details.
*Uit onderzoek van de tanden van de Indohyus blijkt dat het dier herbivoor was en zich bovendien voedde met landplanten , ook al leefde de indohyus al voornamelijk in het water: net zoals nijlpaarden
fig 5
Hatched elements are reconstructed on the basis of related taxa.
Indohyus met een mobieltje ter vergelijking . Het skelet is samengesteld uit botten van meerdere exemplaren van deze dieren, weggespoeld en gefossiliseerd in één aardlaag. (Foto Jeanette Killius) Professor Hans Thewissen, vermoedt dat de dieren ook in het water zijn gaan leven om aan roofdieren te ontkomen. Het Afrikaanse dwerghert (bijvoorbeeld past vandaag nog altijd dezelfde tactiek toe.
( Wikipedia / laatste update = 10 nov 2007 ) De walvissen en evenhoevigen (Cetartiodactyla) zijn een nog niet algemeen geaccepteerde orde van de zoogdieren. Het is een vrij recente samenvoeging van de walvissen en de evenhoevigen. Uit DNA-onderzoek komt regelmatig naar voren dat de walvissen de naaste verwanten van de nijlpaarden zouden zijn. Dit kwam als een grote verrassing omdat op basis van de morfologie en fossiele soorten werd aangenomen dat de walvissen, als afstammelingen van de Mesonychia, de zustergroep van de evenhoevigen zijn en dat beide groepen dus monofyletisch zouden zijn. Bepaalde morfologische kenmerken van uitgestorven, primitieve walvissen, met name van de enkelbeenderen, lijken echter eveneens de plaats van de walvissen binnen de evenhoevigen te ondersteunen, net als modern morfologisch onderzoek van levende soorten. De geldigheid van deze nieuwe indeling is echter nog steeds controversieel.
Genetische gegevens geven echter de volgende verwantschappen:
Eeltpotigen
Varkens en pekari's
Herkauwers
Nijlpaarden
Walvissen
De afstamming van de walvissen ( wikipedia 30 nov 2007 )
Walvissen zijn ontstaan uit landdieren die, waarschijnlijk in het Eoceen, tussen 55 en 34 miljoen jaar geleden, in zee gingen leven. Over de afstamming van de walvis is in de jaren rond de millenniumwisseling veel te doen geweest. In 2001 werden twee 47 miljoen jaar oude en niet complete fossielen ontdekt in de Pakistaanse provincie Beloetsjistan. Ze kregen de namen Pakicetus( Pakicetidae), Rodhocetus balochistanensis en Artiocetus clavis. Het betreft hier tussenvormen tussen op het land levende dieren en de volledig in zee levende walvissen, die nog wel poten hadden. De enkelbeenderen daarvan vertonen kenmerken die anders alleen bij evenhoevigen gezien worden. Er zijn sterke aanwijzingen dat de naaste hedendaagse verwanten de op het land levende nijlpaardenzijn. Vóór de ontdekking van de fossielen had DNA-onderzoek ook al in deze richting gewezen. Men ( dacht tot voor de nieuwe ontdekking ) dat walvissen afstammen van een groep vleesetendeevenhoevigen (Artiodactyla).
( A) Ondertussen is dus de verste bekende verwant van de walvissen dus een dier uit de groep van de land planten-etende INDOHYUS -achtigen , geworden ?
Volgens recente analyses van erfelijk materiaal zijn, binnen de groep van evenhoevigen, de nijlpaarden de nauwste levende verwanten van walvissen. Het gaat een stap te ver om te zeggen dat mijn fossielen die conclusie ondersteunen, aldus de in Utrecht opgeleide Thewissen in een telefonische toelichting. *Want uit de fossielen valt niet op te maken of nu het nijlpaard of een andere evenhoevige de nauwste verwant van de walvissen is. Dat heeft er vooral mee te maken dat er geen heel oude fossielen zijn gevonden van nijlpaarden of nijlpaardachtigen. * De genetische analyse die laat zien dat nijlpaarden de nauwste verwant zijn van dolfijnen en walvissen acht ik niettemin overtuigend."
Planteneters
Lang gingen wetenschappers ervan uit dat walvissen evolueerden uit roofdieren op het land. Dat lag voor de hand, omdat moderne walvissen en dolfijnen geen planteneters zijn. De aanwijzingen dat toch de plantenetende hoefdieren het nauwst verwant zijn aan de walvissen zijn inmiddels echter zeer sterk. Die conclusie is niet alleen gebaseerd op een vergelijking van het DNA. Een extra scharnier in het enkelgewricht zorgt ervoor dat evenhoevigen grotere stappen kunnen nemen dan andere dieren, zegt Thewissen. Dit kenmerk hebben wij ook gevonden in Pakicetusattocki, de vroegste walvis waarvan wij in 2001 het skelet hebben ontdekt.
Graag plaats ik hier ook een kopie van het zeer goede artikel van Tim spaan ( oorspronkelijk te vinden op ) > Na darwin walvis (Tsjok45) )
(1.-) Kan een voor( stam)ouder geologisch jonger zijn , dan zijn nakomelingen ?
Kort antwoord: NEEN . Maar elke stamouder (= common ancestor) heeft minstens twee afstammingslijnen
Uitgebreid genuanceerd antwoord ( van toepassing op het belang van deze vondst )
De wetenschappers hebben hier de skeletten van heel wat levende en dode (zoog)dieren vergeleken om een stamboom te tekenen waarin de walvisachtigen een plaats kregen Elke soort die zij hebben bestudeerd en vergeleken met anderen , is een takje op die boom . Elk takje is verbonden met andere takjes door middelvan een gedeeld voorgeslacht.
The figure shows a consensus cladogram produced by heuristic searches with PAUP (random addition sequence, 1,000 repetitions), using a published data set10. See Supplementary Information for further details.
(Evolutie) Wetenschappers stellen echter soorten nooit op in een aaneengesloten directe ketting van voorvaderen en nakomelingen .
Het is soms mogelijk om het fossiel van één uitgestorven soort te vinden die in een andere uitgestorven soort evolueerde. Maar als de wetenschappers slechts evolutie op die manier zouden bestuderen, dan negeren ze een rijkdom aan andere aanwijzingen over hoe de evolutie opereerde in de geschiedenis van het leven . Aangezien de stamlijnen van zoogdieren evolueeren verwerven ze ( anatomische , morfologische en fossiel traceerbare ) trekken die al de afstammelingen verder uiteen op de stamboom plaatsen .
Ruwweg 150 miljoen jaar geleden evolueerden onze voorvaderen een placenta . (150 million years ago ) De moederkoek wordt gevonden bij mensen, vleermuizen , olifanten, walvissen, en duizenden andere soorten. Niet bij het vogelbekdier (platypus = Ornithorhynchidae (Echte vogelbekdieren)); noch bij Gewone mierenegel (Tachyglossus aculeatus) Platypus en Tachyglossa vertellen ons echter iets over ons voorgeslacht dankzij hun positie op naburige takken van de boom van het leven, . Net als hen, hadden onze eigen voorvaderen ooit GEEN placenta . Dat betekent echter niet, dat de mierenegels of Platypoda (Vogelbekdieren) onze voorvaderen zijn.
Door gelijkaardige methodes te gebruiken, kunnen de wetenschappers Indohyus ,beschouwen als een vertegenwoordiger van een groep die relevante kenmerken vertoont van de de oudste bekende walvissen /walvisachtigen . Indohyus staat ( voorlopig ) het dichtst bij de walvisachtigen.
Volgend diagram ( van carl zimmer ) is gebaseerd op wat de wetenschappers over fossiele walvissen weten en hoe zij( mogelijk ) verwant zijn.
Natuurlijk, is dit slechts een wetenschappelijke hypothese die moet worden getest. En de manier van testen is meer soorten zoals Indohyus ( uit de groep van de raoelliden Raoellidae) te vinden. Paleontologen met wat geluk , zullen meer fossielen moeten zoeken en daaruit waarschijnlijk nieuwe takken bij de basis van de walvisboom kunnen afleiden En (als de huidige hypothese juist is, )zullen heel wat nieuwe soorten Raoliden tot die basis stamlijnen gaan behoren . Die groep kan trouwens ion zijn geheel zowel vóór en/ of als miljoenen jaren na de oudste aquatische walvissen geleefd hebben
Maar dat is niet de kern van de zaak Wat van belang is ,is verwantschap
Misschien beschouwd men het als klein grut of pedeante muggezifterij ; Maar het verwarren van verwanten met voorouders leidt tot misleidende wetenschap , op die manier wordt zeer elegant onderzoek gediskrediteerd . Bovendien is dat een dankbaar handvat voor allerlei anti-wetenschappers en creationisten ....
En, natuurlijk, beweren sommige mensen ( = creationisten )dat het feit dat de verwanten geen directe voorvaderen zijn betekent dat de evolutie zo vals is , als een 11 euro biljet . Maar het is de moeite niet waard aan dergelijke woordspelletjes veel tijd te besteden .... De boven beschreven omschrijvingen bepalen wat de wetenschap als legstukjes van de stamboom puzzel beschouwd ... niet alleen voor walvissen, .... maar ook voor mensen, bloemen, of een ander organisme
(2) zo groot als een "wasbeer " Een ietwat ongelukkige uitdrukking die door de creationisten wordt aangegrepen om nog maar eens te sneren en te proberen paleontologen belachelijk te maken ....
Indohyus heeft het formaat van een wasbeer. ( of van een vos ) Dergelijke vergelijkingen maken wetenschappers om het lekenpubliek een beetje een indruk te geven. Wasberen behoren tot een hele andere orde dan Indohyus (Carnivora en Artiodactyla respectievelijk). Qua anatomie lijkt Indohyus totaal niet op een wasbeer.
(3) " wenkbrauwverdikking " noemt een enkele arrogante creationist het spottend en denigrerend
Het is echter iets helemaal anders Fossiele en moderne walvissen bezitten immers allen een speciaal aangepaste gehoorbeen-structuur ( dikkere mediale( =Involucrum) - & dunnere laterale (tympanic plate) tympanic -wall ) voor een goed onderwater-gehoor ; De dunne "tympanic plate" is trouwens de ( beenderachtige ) walvisversie van het trommelvlies van een landdier
Thewissen : Indohyus heeft dat (= involucrum en die tympanic plate ) ..... " Het is de eerste non- walvis die dat bepaald kenmerk bezit : uiteraard is dat ook het meest spectaculaire en belangrijkste bewijsstuk van diens verwantschap met de wakvissen ."
Toch blijft deze creationist beweren Een wasbeer als voorvader van de walvissen! Maar , dat beweert niemand. Indohyus had het formaat van een wasbeer, niet meer dan dat. Walvissen hebben in dezen niets met wasberen te maken. Niemand beweert dat ook, maar op een één of andere manier denk deze creationist dat de betreffende wetenschappers dat wél beweren. Het is een s
Reacties op bericht (3)
05-11-2008
link eocene whales
<---ffEocene Whales
05-11-2008 om 11:30
geschreven door Tsjok45
25-05-2008
NaDarwin Walvis
Walvis evolutie Over de uitgestorven links in de evolutie van de walvis en hoe deze werden ontdekt.
Een artikel van Tim Spaan.
Verklarende woordenlijst: Mysticeti: baleinwalvissen Odontoceti: tandwalvissen Archaeoceti: oerwalvissen Foerageren: voedsel zoeken Aquatisch dier: zee/waterdier Terrestrisch dier: landdier Amfibisch dier: een in water en op land levend dier
Anno 1859 had Darwin, althans theoretisch, een verklaring hoe de orde der walvissen (Cetacea) was ontstaan. Volgens hem waren walvissen en dolfijnen ontstaan uit landzoogdieren die meer evolutionair voordeel konden halen met het leven en foerageren in het water dan op het land. Dit in tegenstelling tot Lamarck, die in walvissen de aquatische voorouders van landzoogdieren zag. Darwin gaf als voorbeeld een zwarte beer, gezien door een collega in Noord-Amerika, die uren rondzwom in een meertje om insecten te vangen. Als deze beer, of een ander zoogdier, vele generaties lang meer voordeel zou kunnen halen met foerageren in het water of waterrijk gebied, dan op het land, zou dit dier zich vanzelf meer gaan gedragen als waterdier. De betere zwemmers en duikers, die lichamelijk beter waren aangepast aan zwemmen zouden worden bevoorrecht, en langzamerhand zou er een dier ontstaan in de vorm van een zeehond of walvis. Het probleem waar alle argumenten destijds over struikelden was dat er geen bewijs was in de vorm van fossielen. De enige prehistorische walvissen die waren gevonden waren geheel aquatische dieren die Basilosaurus werden genoemd. Een oerwalvis met een 15 meter lang, slangachtig lichaam. Deze waren meer dan eens het slachtoffer van bedrog geweest, een zekere Albert Koch gebruikte de botten van zo'n vijf walvissen om er n groot langgerekt skelet van te construeren dat hij presenteerde als Hydrarchus, de legendarische zeeslang. Paleontoloog Richard Halan ontdekte ook fossielen van Basilosaurus, en hij ging er heel wat wetenschappelijk verantwoordelijker mee om, maar ook hij deed een foute interpretatie. Harlan zag in het dier een reusachtig uitgestorven zeereptiel, dat in lang vervlogen tijden roofzuchtig heerste over de oceanen. Met deze gedachte in het achterhoofd kroonde hij de koning der reptielen: Basilosaurus ('Koningsreptiel'). Pas toen de meest gerespecteerde paleontoloog van Engeland, Sir Richard Owen, een blik wierp op de resten kwam de ware aard van Basilosaurus naar voren. Het gebit van het dier was samengesteld uit smalle tanden voorin de snuit, en achterin brede kiezen ter grote van een aardappel. Dit was in tegenspraak met reptielen, die gewoonlijk een uniform gebit hebben. Een samengesteld gebit met snijtanden, valse kiezen en ware kiezen komt enkel en alleen voor bij zoogdieren. Ook de rugwervels van Basilosaurus waren die van een zoogdier, en kwamen sterk overeen met die van een walvis. De botstructuur onder een microscoop bekeken gaf de doorslag, in plaats van een patroon van buisjes zoals bij reptielen was er onder het cement een patroon met golvende kanaaltjes, zoals van zeezoogdieren. Desondanks bleef Basilosaurus zijn naam van koningsreptiel door taxonomische regeltjes behouden.
Het lange slangachtige lijf van Basilosaurus deed rond 1830 de paleontologen en amateur bottengravers aan zeeslangen denken. Hoewel Basilosaurus tamelijk veel voorkomt in het fossielenbestand werden de korte achterpootjes pas in 1989 gevonden. De lengte van dit dier werd vroeger geschat op wel 45 meter, maar de werkelijke lengte ligt tussen de 15 en 18 meter.
Als oudste en enige bekende Cetacea in het fossielenbestand was Basilosaurus niet bepaald bevorderlijk voor het evolutie-idee. De walvis was geheel aquatisch, en hoewel hij niet zoveel leek op de huidige walvissen (Zoals het gebit, waaruit nou net zijn ware aard was gebleken. Moderne tandwalvissen beschikken over een uniform gebit, van conische pinvormige tanden. Dolfijnen en bruinvissen slikken hun prooi in z'n geheel door, terwijl Basilosaurus en zijn verwanten op hun eten kauwde.) leek hij al helemaal niet op enig bekend landzoogdier. Darwinisten konden alleen afwachten tot er nieuwe vondsten werden gedaan die meer helderheid konden geven. Zoloog William Flower reconstrueerde de landbewonende voorouders van walvissen als: "Primitieve moerasbewonende dieren, haarloos zoals nijlpaarden, maar dan met brede staarten en korte poten, die geleidelijk meer aangepast raakte aan hun omgeving en zo transformeerde tot de huidige walvissen en dolfijnen." Het duurde nog een eeuw voordat de fossielen van de bewuste voorouderlijke dieren werden gevonden, maar tegenwoordig kunnen we zien dat Flowers voorspelling een schot in de roos was.
De door Gingerich gevonden schedel -de witte stukken ontbreken-.
Het lot heeft bepaald dat Phillip Gingerich n van de grootste autoriteiten op het gebied van walvisevolutie is geworden, want toen hij voor het eerst in 1975 Pakistan aandeed om fossielen van primaten te gaan zoeken, waren walvissen niet zijn roeping. Dit stond op het punt te gaan veranderen toen hij met zijn collega's in een 50 miljoen jaar oude rots een paar kleine maar bijzondere schedeldelen opgroef. De eerste indruk was dat het om een behoorlijk onintelligent dier ging, want over de schedel liep een beenkam die gebruikt werd als aanhechting voor de kaakspieren, die maar weinig ruimte overliet voor hersenen. In het laboratorium werden de resten verder onderzocht. Van de oorspronkelijke schedel was niet veel over, alleen het achterste gedeelte, een stuk van de onderkaak en een paar tanden. Te oordelen naar de kop en de tanden zou het kunnen gaan om een Mesonychide.
Mesonychiden zijn betrekkelijk zeldzaam in het fossielenbestand, hun resten worden gevonden in Azi maar zijn meestal fragmentarisch. Wat betreft de constructie van het skelet lijken de Mesonychiden redelijk op elkaar, behalve in grote, daar lopen ze uiteen van Haplodectes, ter grote van een rat, tot Andrewsarchus, met een geschatte lengte van 3,5 meter. Maar wat betreft de vorm van de kiezen of de structuur van de schedel is Haplodectes een miniatuurversie van Andrewsarchus. Mesonychiden zijn geen gemakkelijke dieren voor een paleontoloog om te onderzoeken. Soms lijkt het wel of ze opgebouwd zijn uit verschillende diersoorten. Het waren een soort hoefdieren, maar anders dan alle huidige hoefdieren waren dit vleeseters. Het waren dus roofdieren, maar aan hun tenen hadden ze geen klauwen, in plaats daarvan kleine hoefjes. Deze en andere aanpassingen stelde Mesonychiden in staat hard te rennen. Die snelheid hebben ze klaarblijkelijk niet gebruikt om te jagen, daarvoor komt het slijtagepatroon te weinig overeen met bijvoorbeeld leeuwen. Dus Mesonychiden waren snelle vleesetende hoefdieren, die niet op jacht gingen. De vraag is dus wat ze wel deden. Aaseten lijkt een mogelijkheid. Daarnaast is het idee geopperd dat ze langs stranden zwierven en met hun kaken vissen vingen en schilpadden kraakte. Hoedanook, de paleontologen zijn er nog niet helemaal uit. Wel staat vast dat Mesonychiden de nauwste terrestrische verwanten van de walvissen zijn. Ze komen in vele opzichten overeen met <>Pakicetus, de oudst bekende walvisachtige, en kunnen dus beschouwd worden als vroege (terrestrische) vormen van walvissen.
De Mesonychide Mesonyx
Maar Gingerich ontdekte aan de zijkant van de kop een druifvormig botstukje en een S-vormige beenkrul. Dit sloot alle twijfel over de identiteit van het dier uit, dit kenmerk komt enkel voor bij walvissen, en elke walvis, levend of gefossiliseerd, beschikt erover. Gingerich doopte zijn vondst Pakicetus ('Pakistaanse Walvis'). De bewuste oren waren dan wel die van een walvis, maar toch waren ze bijzonder. Walvissen kunnen onder water de geluidsrichting waarnemen doordat hun oren zijn gevat in een benig kapsel dat, drijvend in een los weefsel van banden en schuim, los ligt van de schedel, zodat ze alleen via de kaak geluiden ontvangen. Pakicetus' druifvormige botjes zaten daarentegen op drie plekken aan de schedel vast. Hij kon onder water wellicht redelijk goed horen, maar er konden nog steeds trillingen door de benige verbindingen sijpelen, waardoor de richting van het geluid werd gemaskeerd. Als walvissen diep duiken vullen ze een netwerk van ruimten rond hun oren met bloed om de oren ook bij hoge druk gesoleerd te houden. De schedel van Pakicetus was in deze gebieden massief, wat suggereert dat hij niet zo diep kon duiken. Dit was een walvis die onder water nog niet zo op zijn gemak was, nog moeite had met horen en zwemmen en die niet in zee maar in ondiepe rivieren leefde. Zowel de meest primitieve als, met vele miljoenen jaren afstand, de oudst bekende walvisachtige. Hierna ging Gingerich actief aan de slag om alle gaten in de walvisevolutie op te vullen. In 1983 en 1985 gingen hij en zijn team naar Egypte om nieuwe opgravingen te verrichten in het zogenoemde 'Dal der Walvissen'. Het 'Dal der Walvissen' dat in feite een kale woestijn is ligt bezaaid met de resten van prehistorische walvissen. Gingerich en zijn team vonden bijna elke dag dat ze er waren een nieuw skelet, het totale aantal gevonden walvissen bedroeg 349. Sommige skeletten waren van Basilosaurus, maar de meeste waren van een kleinere verwant, Dorudon atrox. De reden dat het het Dal der Walvissen zo vol ligt met skeletten van zeedieren is omdat de droge woestijnvlakte tijdens het Eoceen de bodem van een oceaan vormde die paleontologen de Tethyszee noemen. Deze scheidde Afrika van Azi en verbond de Indische en de Atlantische oceaan met elkaar. De Midderlandse Zee is hier een schamel restant van. Het Dal der Walvissen was destijds een beschutte lagune die werd bewoond door haaien, schildpadden, zeekoeien en vele andere zeedieren. Deze lagune was waarschijnlijk een soort kraamkamer voor de kleinere walvissen, soorten zoals Dorudon kwamen hierheen en in de lagune baarden wijfes hun jongen, te oordelen aan de grote hoeveelheden tanden van jonge Dorudons. Alle gevonden Basilosaurirs daarentegen waren volwassen, en deze enorme roofdieren kwamen wellicht alleen naar de lagune om te jagen op de pasgeboren Dorudons. In 1989 startte Gingerich zijn derde expeditie naar Egypte. Dit keer wilde hij de vinnen en heupen van de uitgestorven walvissen eens aan een onderzoek onderwerpen. Aanvankelijk met weinig succes, hij onderzocht gevonden heupen van
De voet en tenen van Basilosaurus. Bron/Source: Gingerich
Basilosaurus, maar deze waren meestal gebroken, onvolledig en vaag. Soms konden onderdelen niet eens gedentificeerd worden. Bij het werk aan een jonge Dorudon vond hij enkele onbekende botten, waarvan n hem deed denken aan een knieschijf. Maar zoiets was ondenkbaar, want als het dier een knieschijf had gehad waren er ook achterpoten, en die zouden voor een volledig aquatisch dier alleen maar lastig zijn. In de laatste week dat ze in Egypte waren ontdekte ze echter nog een nieuw Basilosaurusskelet, waar, 12 meter van de kop verwijderd, een dijbeen werd gevonden. Deze dijbeen werd gevolgd door een knie, een scheen -en kuitbeen, een enkel, en op de laatste dag werden er drie tenen blootgelegd. Een walvis met achterpoten dus, deze spectaculaire ontdekking laat zien dat zelfs in dit volledig aquatische stadium walvissen nog beschikte over kleine restanten van hun vroegere bestaan als landdier. Echter, ondanks dat de rudimentaire achterpootjes van Basilosaurus maar 30 centimeter lang waren, op een lijf van 15 meter, en bevestigt waren aan een bekken dat niet meer in verbinding stond met de ruggengraat (Dus 'zweefde' in het lichaam.) hadden ze nog wl een functie. Gingerich leidde dit af aan de littekens die werden veroorzaakt door spieraanhechtingen, de pootjes beschikte over sterke spieren. Toen Gingerich echter uitprobeerde tot welke bewegingen het pootje in staat was deed er zich weer iets vreemds voor, de pootjes waren erg stijf en onbeweeglijk, want de gladde goot onderaan het dijbeen waar normaliter bij landdieren de knieschijf in glijdt ontbrak. Er waren maar twee standen waarin het been goed paste, gestrekt en tegen het lijf geklemd. Volgens Gingerich zou het been 'knak' gedaan hebben als ze gestrekt of ingetrokken werden. Dit roept uiteraard de vraag op wat het nut hiervan was. Gingerichs eigen hypothese is dat de pootjes een hulpmiddel waren bij de paring, de partners zouden zich aan elkaar kunnen vasthouden. Ze brachten dan hun achterpootjes bij elkaar, deden 'knak' en zaten aan elkaar vast. Ook de kleinere walvis Dorudon beschikte over achterpootjes, maar omdat deze niet over het slangachtige lijf van Basilosaurus beschikte was het duidelijk dat deze lichaamsbouw niet uitsluitend verantwoordelijk was voor de aanwezigheid van de poten. Ze waren in deze fase van de evolutie gewoon nog overgebleven, en verdwenen later weer. Gingerich besefte dat de evolutiepuzzel nog lang niet kompleet was, tussen Pakicetus en Basilosaurus bevond zich nog een gapende leegte van 10 miljoen jaar die opgevuld moest worden. Hierin lag nog een schone taak voor de paleontologen. In 1991 en '92 ging Gingerichs collega Hans Thewissen met zijn team fossielen zoeken in Pakistan, in de hoop om in dit land naast Pakicetus nog meer fossiele walvissen boven water te krijgen. In 1991 ontdekte ze een nieuwe kaak van Pakicetus, completer dan die van Gingerich. Tijdens een tweede expeditie in 1992 ontdekte ze een zoogdier dat leek o
Het skelet van de lopende walvis (Ambulocetus). Bron/source:Whale origins
p een hoefdier of zeekoe. Maar het had wel een raar lijf voor een zoogdier, de voorpoten waren kort, maar met grote platte handen, terwijl de achterpoten eindigden in lange voeten die op clownsschoenen leken. Toen ze de kaak blootlegde kregen ze de driehoekige kiezen in zicht, die worden gevonden in Mesonychiden. Maar net zoals bij Pakicetus werd de identiteit van het dier achterhaald dankzij de oren, toen die tevoorschijn kwamen herkende men meteen het druifvormige oorbotje en de S-vormige beenkrul. Weer had men een vroege walvis gevonden, een lopende walvis dit keer, en een prachtige overgangsvorm. Bij de pas otdekte onderkaak van Pakicetus werd namelijk een klein gaatje ontdekt, zo groot als een speldeknop, aan de binnenkant van de zijvleugel in de onderkaak. Hierdoor lopen bloedvaten en zenuwen naar de kin, en alle landzoogdieren beschikken er over. Bij walvissen is het kleine gaatje echter uitgerekt tot een lange sleuf die langs een groot deel van de kaak loopt. Thewissen zag dat bij het nieuwe fossiel, dat drie miljoen jaar jonger was, het kleine gaatje al wat uitgerekter was, het had een ovaalvorm.
Helaas kon dit spectaculaire fossiel niet in z'n geheel worden meegenomen, want er was een gebrek aan laadruimte. Thewissen moest dus een gedeelte achterlaten, maar kort daarop keerde hij samen met Gingerich terug om ze als nog op te halen en gelijk verder te graven. Zowaar vond Gingerich ook een nieuw skelet, iets jonger dan dat van Thewissen, en meer aquatisch. Een betere zwemmer, want hij werd in gesteenten gevonden die destijds buiten de kust hadden gelegen. In een paar dagen groef hij het op. Het was een dier zo groot als een zeeleeuw, met de kop van een oerwalvis en de tanden van een Mesonychide. Het skelet was tamelijk volledig, al ontbraken de staart en poten, met uitzondering van een dijbeen, die zeer kort zou zijn voor een landdier, maar veel groter dan die van Basilosaurus bijvoorbeeld. Gingerich en Thewissen brachten hun beide vondsten mee naar huis en bestudeerde ze twee jaar lang, tot ze hun onderzoeken in 1994 vlak na elkaar publiceerden. Thewissen noemde zijn vondst Ambulocetus natans, en Gingerich noemde de zijne Rhodocetus kasranii.
Rhodocetus (Dit is het meer komplete skelet dat in 2001 werd gevonden.)
Hierna ging het snel, tussen 1995 en 2001 werden o.a ontdekt: Takracetus simus, Gaviocetus razai, Dalanistes ahmedi, Qaisracetus arifi, Andrewsiphius sloani, Babiacetus indicus, een nieuw species van Basilosaurus (B.drazindai), Basiloterus hussaini enzovoort. Ook werden er meer kompletere skeletten gevonden van Ambulocetus, Rhodocetus (Met poten dit keer.) en Pakicetus. Al spoedig was er geen sprake meer van de gapende leegte in het fossielenbestand die anno1859 heerste wat het walvissen betrof. Men moest nu zelfs soorten gaan afwijzen als voorouders van de huidige walvissen. De familie der Remingtonocetidae bijvoorbeeld, die bestaat uit 5 ontdekte genera waaronder Dalanistes, Remingtonocetus en Kutchicetus, komen niet in aanmerking als directe voorouder van de huidige walvissen. In uiterlijk waren ze hetzelfde als Ambulocetus, maar beschikte over een ongewoon lange en spitse snuit, die veel leek op die van een gaviaal en deze ongetwijfeld op dezelfde manier gebruikte. Kutchicetus bijvoorbeeld zag er ongeveer uit als een otter, maar met de extreem lange snuit van een gaviaal. Zouden je er ooit n in leven zien dan zou het ongetwijfeld het raarste zijn wat je ooit hebt gezien, vergeleken met de 21ste eeuwse zoogdieren.
Deze dieren vormen een deel van de natuur die geheel is verdwenen, hoewel er later nieuwe vormen van amfibische zoogdieren verschenen zoals zeehonden en otters, maar dit waren in feite harige krokodillen, een zoogdiervariant van de Gaviaal of de Alligator. Deze soorten zijn echter verdwenen en alleen de volledig aquatische walvissen hebben het overleefd, wat overigens niet uitsluit dat er misschien nog lang nadat er soorten volledig aquatisch werden nog amfibische soorten zijn gebleven. Het zijn immers twee verschillende ecologische niches, dus beide zaten elkaar niet in de weg. Met de ontdekking van Gaviocetus, wiens heupbeenderen los van de ruggengraat hingen, was de fossiele 'gap' tussen Rhodocetus en Dorudon gedicht. Eindelijk waren alle schakels tussen land en zee gevonden. Echter, vanaf volledig aquatische soorten als Basilosaurus en Dorudon was het evolutieverhaal nog lang niet voorbij. Er stond nog een hoop te gebeuren, want de walvissen zouden zich opsplitsen in twee groepen, de tand -en baleinwalvissen.
De orde der Cetacea wordt opgedeeld in drie subordes; de Odontoceti (Tandwalvissen), de Mysticeti (Baleinwalvissen) en de uitgestorven Archaeoceti ('Oerwalvissen'). Aanvankelijk had men gedacht dat alle drie de ordes zich onafhankelijk van elkaar hadden ontwikkeld uit landzoogdieren, want alle drie de groepen waren zo verschillend van elkaar dat ze bijna onmogelijk een recentelijke gemeenschappelijke voorouder konden hebben, zo leek het. De Archaeo -en Odontoceti hebben beide tanden, maar de eerste hebben zoals al eerder vermeld een gebit dat is samengesteld uit voortanden en knipkiezen, en ze kauwde hun voedsel. De Odontoceti hebben uniforme tanden en slikken prooien in hun geheel door. Ze maken bovendien bovendien gebruik van echolokatie om hun prooi op te sporen. In fossiele resten is de aanwezigheid van echolokatie af te lezen door luchtholten en geluidsweerkaatsende botvlakken, maar bij geen enkele Archaeoceti is zoiets te bespeuren. Bij de Mysticeti weegt elke soort al zo veel als een kudde olifanten, en hebben een bek vol hoornachtige baleinen, die ze gebruiken om kleine vissen en kreeftjes uit het water te zeven. De Mysti -en Odontoceti hebben hun neusgat(en (De Odontoceti hebben er n.)) hoog op hun voorhoofd zitten, waar ze het typische spuitgat vormen, terwijl Archaeoceti hun neus op de gebruikelijk plaats hebben, voorop de snuit.
Toch weet men nu dat de Mysti -en Odontoceti samen zijn voortgekomen uit de Archaeoceti, daar de vroege terrestrische en amfibische walvisachtigen alleen maar Archaeoceti zijn. Maar vooral de manier waarop de baleinwalvissen hieruit zouden zijn ontstaan bleef lange tijd een groot probleem. Eigenlijk had men al in 1966 een fossiele walvis gevonden die de evolutie van oer -naar baleinwalvis laat zien, maar men had hem niet direct als zodanig herkend. Deze walvis, hij heet Aetiocetus, was een tijdlang een buitenbeentje dat niet in de gevestigde ordes paste.
De schedel van Aetiocetus, de baleinwalvis zonder baleinen.
Het was waarschijnlijk geen Archaeoceti, daarvoor was hij te jong en hij had een spuitgat in plaats van neusgaten. Hij beschikte niet over echolocatie, dus een Odontoceti was het ook niet, maar hij had anderhalve centimeter lange tanden dus een Mysticeti was het al helemaal niet. Maar een zekere Leigh Van Valen wist uiteindelijk de vreemde walvis juist te classificeren, het was in alle opzichten een baleinwalvis, alleen met tanden in plaats van baleinen. Een andere bijzondere link tussen oer -en baleinwalvissen is Archaeomysticetus. Deze walvis is 25 miljoen jaar oud en was ongeveer 9 meter lang, hij had een kop van 1,70 meter. Hij beschikte over wat kenmerken van de Mysticeti zoals twee losse onderkaakhelften, en de bovenkaak begon achter het oog door te lopen (Waarbij het spuitgat naar achteren werd getrokken tot op het voorhoofd.), maar zijn tanden zouden zo uit de kaak van Basilosaurus kunnen komen. Ze waren ook even groot als die van Basilosaurus, en hadden dezelfde onderlinge afstand. Baleinwalvissen begonnen oorspronkelijk dus als roofwalvissen, die vissen en haaien met hu tanden grepen en inslikten. Maar een school kril uit het water zeven konden ze nog niet. Hoe baleinwalvissen hun techniek van voedsel uit het water zeven, en de daarvoor benodigde baleinen ontwikkelden kunnen we niet op de voet volgen. Baleinen fossiliseren moeizaam, de oudste is 15 miljoen jaar oud, veel later dan de eerste baleinwalvissen ontstonden. Er is misschien een mogelijkheid die wordt ondersteund door het fossielenbestand en embryologisch onderzoek. De baleinwalvis Llanocetus denticrenatus leefde 34 miljoen jaar geleden. Voorin zijn bek beschikte hij over de pinvormige voortanden van de Archaeoceten, en achterin zijn bek stonden een soort stervormige kiezen. Mogelijk is het zeven van voedsel begonnen met die kiezen, want vandaag de dag is er een diersoort met vergelijkbare kiezen die deze gebruikt om kril uit het water te zeven, het is de zeehond 'krabeter'. Llanocetus verzamelde zijn voedsel waarschijnlijk met een combinatie van jagen en zeven, hij gebruikte zijn voortanden om snelle vissen te grijpen, en waarneer hij een school krill tegenkwam verslond hij daar een bek vol van en liet het water weer tussen zijn kiezen vandaan lopen. In baleinwalvisembryo's valt bovendien af te lezen dat op een bepaald moment, vlak voor de tanden geheel verdwijnen, ze eerst kleiner en nvormiger worden en fors in aantal toenemen, zodat de hele bek vol komt te staan. Wellicht werd er een woud van dunne tanden gevormd die als zeef dienstdeden? Zoals het er nu uitziet maakte walvissen in eerste instantie dus gebruik van hun tanden en kiezen om water te zeven, waarna ze overstapten op baleinen. Hoe de baleinen zelf zijn ontstaan is dus nog onbekend, hoewel ze mogelijk zijn voortgekomen uit tandvleesplaten. De huidige Dall-bruinvis is een tandwalvis, maar zijn bek wordt geheel gedomineerd door hoornachtige tandvleesplaten die hij gebruikt om prooi te grijpen. Microscopisch gezien verschillen ze in structuur niks van baleinen. Wellicht ging 30 miljoen jaar geleden een voorouder van de baleinwalvissen dezelfde kant op, en ontwikkelde hoornachtige baleinplaten aan zijn tandvleesranden waarmee water en plankton effectief konden worden gescheiden.
De oorsprong van de tandwalvissen is misschien nog wel lastiger. Niet zozeer omtrent het verschillende gebit en de plaatsing van de neusgaten, net als vroege baleinwalvissen beschikken ook vroege tandwalvissen, bijvoorbeeld Squalodon, over de driehoekige kiezen die ze van hun Archaeocetische voorouders erfden, en stond hun neus iets meer naar achteren, als wel om de vorming van hun echolokatiesysteem.
De tandwalvis Squalodon beschikte nog over de driehoekige kiezen van zijn voorouders zoals Dorudon en Basilosaurus.
Voor het ontstaan van het echolocatie zijn helaas geen bewijzen, enkel wat plausibele hypothesen. De oren van tandwalvissen waren 10 miljoen jaar voor de eerste tandwalvis al gedeeltelijk van de schedel losgekomen, zodat de latere, met echolocatie uitgeruste walvissen voor hun eigen klikken waren afgeschermd. Voor hun nieuwe geluiden moesten ze leren om met hun neus geluid te maken in plaats van met de stembanden. Verschillende zoogdierkenners hebben bevestigd dat dit al wordt gedaan door steenbokken, gemzen en gazellen. Die slaken soms met hun neus alarmkreten, en wellicht deden de Mesonychiden dat eveneens. Het meloenorgaan is waarschijnlijk ontstaan als een soort neusstop. Een keer ontdekte men bij het ontleden van een baleinwalvis bij het spuitgat een paar kleine klontjes vet en weefsel die er uitzagen als miniatuurmeloentjes. Men denkt nu dat het meloenorgaan is ontstaan bij Archaeoceti, die ze gebruikte om bij het duiken de neusgaten af te sluiten en zout water tegen te houden. Bij baleinwalvissen zijn ze vervolgens gekrompen en hebben hun functie verloren, terwijl bij tandwalvissen er n opzwol en een nieuwe functie kreeg. Hiermee beschikte de vroege tandwalvissen over de basismiddelen voor echolocatie. De grote lijnen van walvisevolutie liepen hiermee ook op hun eind, na het ontstaan van de Mysti -en Odontoceti ontstonden er verschillende variaties, zoals vinvissen ten opzichte van ware walvissen, en verschillende families dolfijnen, bruinvissen en grote tandwalvissen als zoals de potvis. Maar echt grote transformaties werden er niet meer ondergaan, de Cetacae waren volkomen aangepast aan het waterleven.
In het kort nog een keer; de chronologische geschiedenis:
Mesonychiden
Tijd: 60 - 32 miljoen jaar geleden Terrestrische en vleesetende hoefdieren.
Pakicetidae
Tijd: 53 - 50 miljoen jaar geleden 3 genera bekend, waaronder Pakicetus en Ichthyolestes. De oudste Cetacea, wat betreft anatomie volledig terrestrische walvissen.
Ambulocetidae
Tijd: 49 miljoen jaar geleden 3 genera waaronder Ambulocetus en Himalayacetus, amfibische, krokodilachtige walvissen.
Remingtonocetidae
Tijd: 49-43 miljoen jaar geleden 5 genera waaronder Dalanistes en Kutchicetus. Ongeveer gelijk als Ambulocetiden, maar met langere schedels en andere kenmerken. Waarschijnlijk vroege vormen, maar geen directe voorouders van huidige walvissen.
Protocetidae
Tijd: 49-46 miljoen jaar geleden 9 genera waaronder Rhodocetus, Protoceiden worden enkel gevonden in lagen die ooit de bodem van kustwateren vormden. Ze leefde dus al buiten rivieren en meren, maar bleven in de buurt van de kust.
Basilosauridae
Tijd: 40-35 miljoen jaar geleden 4 genera waaronder Basilosaurus, Dorudon en Zygorhiza (hier afgebeeld). Volledig aquatische walvissen, en als gevolg hiervan worden ze opeens op veel grotere afstand gevonden vergeleken met de voorgaande soorten, die allemaal rond Pakistan en India zijn gevonden. Basilosauriden zijn gevonden in Egypte, Engeland, en de langs de Westkust van Noord-Amerika.
Odonto -en Mysticeti
Tijd: 34 miljoen jaar geleden t/m heden Tand -en baleinwalvissen, vroege soorten tonen sterke overeenkomsten met Basilosauridae, later gaan beiden een eigen richting op. Mysticeti ontwikkelen baleinen om kleine prooi uit het water te zeven. Meer dan 130 moderne en fossiele genera overigens.
Het terugwijkende spuitgat. De rode pijl geeft de plaats van de neusgaten aan. Weergegeven fossielen: Pakicetus - 50 miljoen jaar oud. Zygorhiza - 35-30 miljoen jaar oud. Squalodon - 28-15 miljoen jaar oud.
Bronnen:
1 Aan de waterkant, Carl Zimmer 1999 De daaruit voortgekomen feiten worden bevestigd door primaire literatuur van Gingerich, Thewissen, Uhen, Fish, enz.
Although the first ten million years of whale evolution are documented by a remarkable series of fossil skeletons, the link to the ancestor of cetaceans has been missing. It was known that whales are related to even-toed ungulates (artiodactyls), but until now no artiodactyls were morphologically close to early whales. Here we show that the Eocene south Asian raoellid artiodactyls are the sister group to whales. The raoellid Indohyus is similar to whales, and unlike other artiodactyls, in the structure of its ears and premolars, in the density of its limb bones and in the stable-oxygen-isotope composition of its teeth. We also show that a major dietary change occurred during the transition from artiodactyls to whales and that raoellids were aquatic waders. This indicates that aquatic life in this lineage occurred before the origin of the order Cetacea.
Phylogenetic analyses of molecular data on extant animals strongly support the notion that hippopotamids are the closest relatives of cetaceans (whales, dolphins and porpoises)1, 2, 3. In spite of this, it is unlikely that the two groups are closely related when extant and extinct artiodactyls are analysed, for the simple reason that cetaceans originated about 50 million years (Myr) ago in south Asia, whereas the family Hippopotamidae is only 15 Myr old, and the first hippopotamids to be recorded in Asia are only 6 Myr old4. However, analyses of fossil clades have not resolved the issue of cetacean relations. Proposed sister groups ranged from the entire artiodactyl order5, 6, to the extinct early ungulates mesonychians7, to an anthracotheroid clade8 (which included hippopotamids), to weakly supporting hippopotamids (to the exclusion of anthracotheres9, 10).
The middle Eocene artiodactyl family Raoellidae11, 12, 13, 14 is broadly coeval with the earliest cetaceans, and both are endemic to south Asia. Raoellids, as a composite consisting of several genera, have been added to some phylogenetic analyses5, 10, but no close relation to whales was found because raoellid fossils were essentially limited to dental material11, 12, 13, 14. We studied new dental, cranial and postcranial material for Indohyus, a middle Eocene raoellid artiodactyl from Kashmir, India (Fig. 1). All fossils of Indohyus were collected at a middle Eocene bone bed extending for about 50 m at the locality Sindkhatudi in the Kalakot region of Kashmir on the Indian side of the Line of Control. Our analysis identifies raoellids as the sister group to cetaceans and bridges the morphological divide that separated early cetaceans from artiodacyls. This has profound implications for the character transformations near the origin of cetaceans and the cladistic definition of Cetacea, and identifies the habitat in which whales originated. Taken together, our findings lead us to propose a new hypothesis for the origin of whales.
a, b, Oblique lateral view of skull RR 208 (a) and ventral view of skull RR 207 (b). ch, Posterior views of humerus (RR 149, c) and femur (RR 101, d), plantar views of metacarpal (RR 138, e) and proximal manual phalanx (RR 19, f), dorsal view of astragalus (RR 224, g), and posterior view of metatarsal (RR 139, h). il, Histological mid-shaft sections for humerus of the pakicetid Ichthyolestes (H-GSP 96227, i), humerus of Indohyus (RR 157, j), femur of Indohyus (RR 42, k) and femur of the artiodactyl Cainotherium (IVAU unnum, l). Both scale bars are 1 cm; the scale bar near d goes with ah, and that near l goes with il.
To investigate the importance of raoellids in cetacean phylogeny, we excluded raoellids from our initial phylogenetic analysis of artiodactyls plus cetaceans. Our data set differed from previous analyses10 by the addition of several archaic anthracotheres, and some corrected scores for pakicetid cetaceans. This analysis found stronger support for hippopotamidcetacean sister-group relations than the previous analysis10, consistent with molecular studies1, 2, 3. However, the base of the artiodactyl cladogram is poorly resolved (see Supplementary Information for details on phylogenetic runs). In a second cladistic analysis (Fig. 2), we added the raoellids Khirtharia and Indohyus as well as several archaic ungulate groups (condylarths) and found that raoellids and cetaceans are sister groups and that they are the basal node in the Cetacea/Artiodactyla clade, consistent with some previous analyses that used different character sets5, 6. Our analysis is the first to show that raoellids are the sister group to cetaceans, resolving the biogeographic conundrum and closing the temporal gap between cetaceans and their sister. Relations between most artiodactyl families higher in the tree are poorly resolved, and our data lack implications for the relations between these families. Our analysis strongly argues that raoellids and cetaceans are more closely related to each other than either is to hippopotamids.
The figure shows a consensus cladogram produced by heuristic searches with PAUP (random addition sequence, 1,000 repetitions), using a published data set10. See Supplementary Information for further details.
Indohyus shares with cetaceans several synapomorphies that are not present in other artiodactyls. Most significantly, Indohyus has a thickened medial lip of its auditory bulla, the involucrum (Figs 1 and 3), a feature previously thought to be present exclusively in cetaceans. Involucrum size varies among cetaceans, but the relative thickness of medial and lateral walls of the tympanic of Indohyus is clearly within the range of that of cetaceans and is well outside the range of other cetartiodactyls (Fig. 3). Other significant derived similarities between Indohyus and cetaceans include the anteroposterior arrangement of incisors in the jaw, and the high crowns in the posterior premolars.
In cetaceans (open squares), the medial tympanic wall is inflated and called the involucrum, and the lateral tympanic wall is thinned and called the tympanic plate. In artiodactyls (open triangles), the medial and lateral tympanic walls are more similar in thickness, causing values on the y axis to be closer to 1. See Supplementary Information for further details.
Until now, the involucrum was the only character occurring in all fossil and recent cetaceans but in no other mammals5, 15, 16. Identification of the involucrum in Indohyus calls into question what it is to be a cetacean: it requires either that the concept of Cetacea be expanded to include Indohyus or that the involucrum cease to characterize cetaceans. We argue that the content of Cetacea should remain stable and include Pakicetidae, Ambulocetidae, Remingtonocetidae, Protocetidae, Basilosauridae, Mysticeti and Odontoceti6, 7, 9, 10, 17. Thus, Cetacea remains a monophyletic group, whereas Artiodactyla remains a paraphyletic group (because Raoellidae are included but Cetacea are excluded). An alternative classification would render both Cetacea and Artiodactyla monophyletic by including Raoellidae in Cetacea and by limiting Artiodactyla to those clades one node above the raoellid/cetacean node (Fig. 2). We do not prefer this classification because it causes instability by significantly altering the traditional content of both Artiodactyla and Cetacea.
Characters identified as synapomorphies for Cetacea in some of our most parsimonious trees include: long external auditory meatus, double-rooted P3/, lack of P4/ protocone, M1-2/ metacones present but small, and lack of M1-2/ hypocone. None of these features characterize all modern and extinct cetaceans; the dental characters, for instance, cannot be scored in toothless mysticetes. In addition, all of these characters are found in some mammals unrelated to cetaceans.
We attach particular importance to two character complexes that characterize basal cetaceans, constitute synapomorphic suites for the order, and are of great functional importance. All fossil and recent cetaceans differ from most other mammals in the reduction of crushing basins on their teeth: there are no trigonid and talonid basins in the lower molars, and the trigon basin of the upper molars is very small (for example in pakicetids and ambulocetids) or absent. Crushing basins are large in raoellids (Fig. 1a, b) and other basal ungulates. This implies that a major change in dental function occurred at the origin of cetaceans, probably related to dietary change at the origin18. Reduced crushing basins also occur in mesonychids, archaic ungulates long thought to be closely related to cetaceans. However, mesonychian molars have wear facets very unlike those of cetaceans7, 18, whereas wear facets in raoellids are more similar to wear facets in early cetaceans14.
The second character complex that identifies cetaceans is the shape of the postorbital and temporal region of the skull. In early cetaceans, this region is long and narrow19. This affects the sense organs: the olfactory peduncle is long and narrow and the orbits are set close together near the roof of the skull. It also affects oral function, the nasopharyngeal duct is narrow, and the out-lever of the masticatory muscles is long, increasing the closing speed of the jaws. We speculate that the importance of different sense organs was related to these changes, or that changing diet led to a change in food-processing organs.
Behaviourally, the earliest whales (pakicetids) were aquatic waders5, 20, 21, 22, 23. This led us to investigate whether Indohyus was aquatic too. Cortical bone thickness in secondarily aquatic tetrapods is commonly increased at the expense of the medullary cavity, a pattern called osteosclerosis24. Osteosclerosis occurs in early whales20, 21, 25, manatees26, 27, sea otters28, Hippopotamus29, beavers29, pinnipeds29 and Mesozoic marine reptiles26, 30. Osteosclerosis provides ballast that allows some aquatic taxa to be bottom walkers (hippopotamids) and others to maintain neutral buoyancy in water (manatees)30. Histological sections indicate that the limb bones of Indohyus are also osteosclerotic (Fig. 1il), in a similar manner to those of pakicetid cetaceans. Our survey of cortical bone thickness in the limb bones of terrestrial artiodactyls shows that this pattern is unusual for that order: in mid-bone cross-sections of the femur, the medullary cavity makes up between 0.60 and 0.75 of the width of the bone, whereas in aquatic mammals the values are lower (Hippopotamus, 0.55; pakicetids and ambulocetids, 0.250.57). In Indohyus this ratio is 0.42, suggesting that Indohyus was osteosclerotic and thus aquatic. We interpret the limb osteosclerosis of Indohyus to be related to bottom walking and not to slow swimming, because the limbs are gracile and not modified into paddles.
To investigate further the hypothesis that Indohyus was aquatic, we studied the stable isotopes of its enamel, a tissue relatively resistant to preburial and postburial alteration of isotopic composition31. Enamel 18O values are influenced by the oxygen isotope composition of the food and water ingested by an animal as well as by certain physiological processes (such as sweating, panting and respiration)32. For aquatic species, the flux of environmental water by means of direct ingestion and transcutaneous exchange overwhelms all other oxygen sources33 and can cause the enamel 18O values of freshwater taxa (for example Hippopotamus) to be 23 lower than those for terrestrial mammals33, 34. Mean 18O values for four individuals of Indohyus are at least 2 lower than those for our comparative sample of Eocene terrestrial and semi-aquatic mammals from formations of India and Pakistan of similar or slightly older age (Fig. 4)22, 23. Although not representative of the specific deposits from which Indohyus was collected, oxygen isotope values for each ecological type from these sites (namely terrestrial, 2428; semi-aquatic, 23 or less) are surprisingly consistent regardless of age or location (Fig. 4). This suggests that temporal and spatial variation in environmental isotope values was relatively minor and was most probably insufficient to account for the extremely low 18O we have reported for Indohyus. We did not recover tooth material of other mammals at the Indohyus site; until such material can be analysed, the most consistent interpretation is that these low values are a result of the aquatic habits of this species.
Results are shown as means s.d. for the sample population. Filled triangle, Indohyus; open triangle, Khirtharia; open squares, terrestrial mammals; filled squares, brackish-water anthracobunids; filled circles, freshwater/brackish-water archaeocetes. See Supplementary Information for details.
Supporting evidence for a semi-aquatic life of Indohyus comes from examination of its inter-individual variation in 18O values. The overwhelming influx and mitigating influence of isotopically homogeneous environmental water causes the variation in individual 18O values for semi-aquatic and aquatic species (s.d. < 0.5) to be much lower than that of terrestrial species (s.d. > 1.0)33. This line of evidence is especially relevant in our study because it does not require an approximation of the mean environmental isotope values for a site through analysis of the associated fauna. Variation in 18O values for Indohyus (s.d. = 0.4) is extremely low and when compared with species of sufficient sample size (n > 3) it is similar to that of the semi-aquatic archaeocete Pakicetus. Given that the influence of physiological and environmental factors on body water 18O values is more strongly felt at smaller body sizes35, this low level of variation is particularly compelling for Indohyus, with a body mass of less than 50 kg.
To explore the diet of Indohyus we studied carbon isotopes. Enamel 13C values are defined by the carbon isotope composition of an animal's diet and can be used to identify the food webs and resources used by an animal36. The 13C values of primary producers at the base of aquatic and terrestrial food webs overlap, but values for freshwater phytoplankton are typically depleted in 13C relative to freshwater macrophytes37, and both types of aquatic producer are depleted in 13C relative to terrestrial C3 plants37, 38, 39. Consumers foraging within food webs fuelled by freshwater phytoplankton (for example freshwater and brackish-water foraging Eocene whales) typically have lower 13C values than species foraging on aquatic macrophytes37 or on terrestrial resources33 (Fig. 4). Enamel 13C values for Indohyus are higher than those for most early cetaceans and are most similar to the 13C values in enamel for terrestrial mammals from early and middle Eocene deposits in India and Pakistan. Indohyus could have been feeding on land or in water, but it was clearly eating something different from archaeocetes such as Pakicetus and Ambulocetus. If the large crushing basins in the molars of Indohyus were used for processing vegetation, these 13C values in enamel could come from the ingestion of terrestrial plants or aquatic macrophytes. Alternatively, a more ominivorous diet would suggest that Indohyus might have foraged on benthic, aquatic invertebrates in freshwater systems. Although we cannot exclude the possibility of aquatic foraging by Indohyus, 13C values in enamel do suggest that the diet of Indohyus differed significantly from that of Eocene whales. A more refined interpretation of the dietary preferences of Indohyus will require a study of tooth wear and tooth morphology.
Indohyus was a small, stocky artiodactyl, roughly the size of the raccoon Procyon lotor (Fig. 5). It was not an adept swimmer; instead it waded in shallow water, with its heavy bones providing ballast to keep its feet anchored. Indohyus may have fed on land, although a specialized aquatic diet is also possible.
The modern artiodactyl morphologically most similar to Indohyus is probably the African mousedeer Hyemoschus aquaticus. Hyemoschus lives near streams and feeds on land, but flees into the water when danger occurs40. Indohyus had more pronounced aquatic specializations than Hyemoschus does, and it probably spent a considerably greater amount of time in the water either for protection or when feeding. As indicated by the evidence from stable isotopes, Indohyus spent most of its time in the water and either came onshore to feed on vegetation (as the modern Hippopotamus does) or foraged on invertebrates or aquatic vegetation in the same way that the modern muskrat Ondatra does.
Raoellids are the sister group to cetaceans, and this implies that aquatic habitats originated before the Order Cetacea. The great evolutionary change that occurred at the origin of cetaceans is thus not the adoption of an aquatic lifestyle. Here we propose that dietary change was the event that defined cetacean origins; this is consistent with the cranial and dental synapomorphies identified. Molars of Indohyus are markedly different from those of pakicetids, and it is widely assumed that pakicetids ate aquatic prey18, 23.
Our working hypothesis for the origin of whales is that raoellid ancestors, although herbivores or omnivores on land, took to fresh water in times of danger. Aquatic habits were increased in Indohyus (as suggested by osteosclerosis and oxygen isotopes), although it did not necessarily have an aquatic diet (as suggested by carbon isotopes). Cetaceans originated from an Indohyus-like ancestor and switched to a diet of aquatic prey. Significant changes in the morphology of the teeth, the oral skeleton and the sense organs made cetaceans different from their ancestors and unique among mammals.
We chose an existing character matrix10 as the basis for our phylogenetic analysis. We corrected scores of some of the taxa, and made some changes in the taxa included. Details on these taxa, the rationale for using them, and their scores are given in Methods and in Supplementary Table 1.
Tympanic wall thickness was investigated to address the presence of the involucrum quantitatively. We calculated the ratio of medial tympanic wall thickness divided by lateral tympanic wall thickness (see Methods and Supplementary Table 2). Bone histology was studied to investigate the presence of osteosclerosis. It was quantified as the ratio of medullary cavity width divided by bone width in the mediolateral plane (see Methods and Supplementary Table 3).
For analysis of stable isotopes, we prepared powders by following published methods (see Methods and Supplementary Tables 4 and 5). Multiple samples were collected for each species to provide an estimate of population means for carbon and oxygen isotope values33. About 5 mg of enamel powder was collected from each specimen (tooth) for study of carbon and oxygen isotope values.
Most fossils of Indohyus were collected by the late Indian geologist A. Ranga Rao, who discovered the locality about 25 years ago (acronym RR); additional fossils were collected by S.B. and B.N.T. at the same locality (acronym IITR-SB-Kal-S).
Milinkovitch, M. C., Bérubé, M. & Palsbøl, P. J. in The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea (ed. Thewissen, J. G. M.) 113131 (Plenum, New York, 1998)
Nikaido, M., Rooney, A. P. & Okada, N. Phylogenetic relationships among cetartiodactyls based on insertions of short and long interspersed elements: Hippopotamuses are the closest extant relatives of whales. Proc. Natl Acad. Sci. USA96, 1026110266 (1999) | Article | PubMed | ChemPort |
Gatesy, J. & O'Leary, M. A. Deciphering whale origins with molecules and fossils. Trends Ecol. Evol.16, 562570 (2001) | Article |
Boisserie, J.-R., Lihoreau, F. & Brunet, M. Origins of Hippopotamidae (Mammalia, Cetartiodactyla): towards resolution. Zool. Scr.34, 119143 (2005) | Article |
Thewissen, J. G. M., Williams, E. M., Roe, L. J. & Hussain, S. T. Skeletons of terrestrial cetaceans and the relationships of whales to artiodactyls. Nature413, 277281 (2001) | Article | PubMed | ISI | ChemPort |
Theodor, J. M. & Foss, S. E. Deciduous dentitions of Eocene cebochoerid artiodactyls and cetartiodactyl relationships. J. Mammal. Evol.12, 161181 (2005) | Article |
O'Leary, M. A. in The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea (ed. Thewissen, J. G. M.) 133161 (Plenum, New York, 1998)
Boisserie, J.-R., Lihoreau, F. & Brunet, M. The position of Hippopotamidae within Cetartiodactyla. Proc. Natl Acad. Sci. USA102, 15371541 (2005) | Article | PubMed | ChemPort |
Geisler, J. H. & Uhen, M. D. Morphological support for a close relationship between hippos and whales. J. Vertebr. Paleontol.23, 991996 (2003) | Article |
Geisler, J. H. & Uhen, M. D. Phylogenetic relationships of extinct Cetartiodactyls: results of simultaneous analyses of molecular, morphological, and stratigraphic data. J. Mammal. Evol.12, 145160 (2005) | Article |
Ranga Rao, A. New mammals from Murree (Kalakot zone) of the foot-hills near Kalakot, J & K State. J. Geol. Soc. India12, 125134 (1971)
Ranga Rao, A. & Misra, V. N. On the new Eocene mammal localities in the Himalayan foot-hills. Himalayan Geol.11, 422428 (1981)
Kumar, K. & Sahni, A. Eocene mammals from the Upper Subathu Group, Kashmir Himalaya, India. J. Vertebr. Paleontol.5, 153168 (1985)
Thewissen, J. G. M., Williams, E. M. & Hussain, S. T. Eocene mammal faunas from northern Indo-Pakistan. J. Vertebr. Paleontol.21, 347366 (2001) | Article |
Luo, Z. in The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea (ed. Thewissen, J. G. M.) 269301 (Plenum, New York, 1998)
Luo, Z. & Gingerich, P. D. Terrestrial Mesonychia to aquatic Cetacea: transformation of the basicranium and evolution of hearing in whales. Univ. Mich. Pap. Paleontol.31, 198 (1999)
Thewissen, J. G. M & Williams, E. M. The early radiations of Cetacea (Mammalia): evolutionary pattern and developmental correlations. Annu. Rev. Ecol. Syst.33, 7390 (2002) | Article | ISI |
O'Leary, M. A. & Uhen, M. D. The time of origin of whales and the role of behavioral changes in the terrestrialaquatic transition. Paleobiology25, 534556 (1999)
Nummela, S., Hussain, S. T. & Thewissen, J. G. M. Cranial anatomy of Pakicetidae (Cetacea, Mammalia). J. Vertebr. Paleontol.26, 746759 (2006) | Article |
Gray, N. M., Kainec, K., Madar, S., Tomko, L. & Wolfe, S. Sink or swim? Bone density as a mechanism for buoyancy control in early cetaceans. Anat. Rec.: Adv. Int. Anat. Evol. Biol.290, 638653 (2007) | Article |
Madar, S. I. The postcranial skeleton of early Eocene pakicetid cetaceans. J. Paleontol.81, 176200 (2007) | Article |
Roe, L. J. et al. in The Emergence of Whales: Evolutionary Patterns in the Origin of Cetacea (ed. Thewissen, J. G. M.) 399422 (Plenum, New York, 1998)
Clementz, M. T., Goswami, A., Gingerich, P. D. & Koch, P. L. Isotopic records from early whales and sea cows: contrasting patterns of ecological transition. J. Vertebr. Paleontol.26, 355370 (2006) | Article |
Francillon-Vieillot, H. et al. in Skeletal Biomineralization Patterns, Processes, and Evolutionary Trends (ed. Carter, J. G.) 471530 (Van Nostrand Reinhold, New York, 1990)
de Buffrénil, V., Ricqlès, A., Ray, C. E. & Domning, D. P. Bone histology of the ribs of the archaeocetes (Mammalia: Cetacea). J. Vertebr. Paleontol.10, 455466 (1990)
Kaiser, H. E. Untersuchungen zur vergleichenden Osteologie der Fossilen und rezenten Pachyostosen. Palaeontographica A114, 113196 (1960)
Domning, D. P. & de Buffrénil, V. Hydrostasis in the Sirenia: quantitative data and functional interpretations. Mar. Mamm. Sci.7, 331368 (1991) | Article | ISI |
Fish, F. E. & Stein, B. R. Functional correlates of differences in bone density among terrestrial and aquatic genera in the family Mustelidae (Mammalia). Zoomorphology110, 339345 (1991) | Article |
Wall, W. P. The correlation between high limb-bone density and aquatic habitats in recent mammals. J. Paleontol.57, 197207 (1983)
Taylor, M. A. Functional significance of bone ballast in the evolution of buoyancy control strategies by aquatic tetrapods. Histor. Biol14, 1531 (2000)
Koch, P. L., Tuross, N. & Fogel, M. L. The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hydroxyapatite. J. Archaeol. Sci.24, 417429 (1997) | Article |
Kohn, M. J. Predicting animal 18O: accounting for diet and physiological adaptation. Geochim. Cosmochim. Acta60, 48114829 (1996) | Article | ChemPort |
Clementz, M. T. & Koch, P. L. Differentiating aquatic mammal habitat and foraging ecology with stable isotopes in tooth enamel. Oecologia129, 461472 (2001)
Bocherens, H., Koch, P. L., Mariotti, A., Geraads, D. & Jaeger, J.-J. Isotopic biogeochemistry (13C, 18O) of mammalian enamel from African Pleistocene hominid sites. Palaios11, 306318 (1996) | Article | ISI |
Bryant, J. D. & Froelich, P. N. A model of oxygen isotope fractionation in body water of large mammals. Geochim. Cosmochim. Acta59, 45234537 (1995) | Article | ChemPort |
Jim, S., Ambrose, S. H. & Evershed, R. P. Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen and apatite: Implications for their use in palaeodietary reconstruction. Geochim. Cosmochim. Acta68, 6172 (2004) | Article |
Is ID de wetenschappelijke (sic) uitleg voor creationiisten 150jaar Darwinisme... De evolutietheorie gewikt en gewogen David sorensen en revolutietheorie lalala DE VERRIJZENIS VAN DE JONGE VERDRONKEN KOE » Reageer (21) ID TOVERDOOS OVER SLECHTE ID-EETJES ,RESISTENTIE & TOVERDOCTORS c) OEC d) YEC
Een oeroud spoor Denisova : ZUID SIBERIË DENISOVA - mens
DMANISI AAP OF MENS DE OUDSTE ? » Reageer (1)Nakalipithecus nakayamai (<) Alweer eentje ? ..... De tand des tijds / Heidelberg-mens DE HEUPEN VAN EVA CASABLANCA MAN /erectus Hobbit is aparte soort ? » Reageer (2) NOG EEN BENDE BIJTERS POLONAISE met neanderthaler TANDEN UIT DE QESEM GROT Neanderthaler genoom Mitochondriale Genenkaart van Neanderthaler Xuchang mens
b)Biologie 1.-(EVOLUTIE ) NAS / IM Document 2008 (2) NAS / IM Document 2008 ( 1)
Cambrium & precambrium AVALON GABONESE chips
Evolutie in actie Opmerkelijk snelle adaptaties bij kroatische ruine-hagedissen:
EEN PLUIM VOOR CHINA Geef eens een pootje Pluimgewicht KLEIN DUIMPJE & DE REUS &VEREN Opnieuw gevederde Maniraptor Similicaudipteryx. Aerosteon riocoloradensis
Krokodillen
PAKASUCHUS KAPILIMAI Prestosuchus chiniquensis Terug naar zee viseter
Eritherium azzouzorum LJOEBA Darwinius masillae VLIEGENDE KATTEN ? Vleermuizen ALWEER EEN BELANGRIJK STUK UIT DE LEGPUZZEL Zee-zoogdieren ; Van de wal in de visgronden Indohyus / walvisevolutie Zeehonden-evolutie
*Blauwe en groene ( =nederlandse)teksten zijn meestal aanklikbare links *Engelse Wikipedia teksten verwijzen in de linkerkolom naar verschillende niet-engelse versies van het wikiartikel *Blauwe teksten tusssen "" , zijn voornamelijk ( gedeeltelijke)citaten afkomstig van mensen met andere meningen
Evolutie / Charles Darwin aan de basis: SELECTIE NATUURLIJK DE LEVENSBOOM UITSTERVEN DEEP TIME Biogeografie SEKSUELE SELECTIE Al jaren lang samen CO-EVOLUTIE De economie van de natuur Geleidelijke veranderingen
Unintelligent design:
IP Theorie Ons gebit , De appendix, Geboorte, De weg van het zaad , Nervus Laryngeus Recurrens, DNA , 75 mistakes , Het boek
Unintelligent design (1): Rechtop lopen Tomaso over het oog , oogspieren , Unintelligent Design (5): ZWEETVOETEN Unintelligent design (6): Vitamine C Unintelligent design (8): Kuitspieren
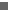






 s.d. for the sample population. Filled triangle, Indohyus; open triangle, Khirtharia; open squares, terrestrial mammals; filled squares, brackish-water anthracobunids; filled circles, freshwater/brackish-water archaeocetes. See
s.d. for the sample population. Filled triangle, Indohyus; open triangle, Khirtharia; open squares, terrestrial mammals; filled squares, brackish-water anthracobunids; filled circles, freshwater/brackish-water archaeocetes. See 









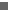


















 18O values are influenced by the oxygen isotope composition of the food and water ingested by an animal as well as by certain physiological processes (such as sweating, panting and respiration)
18O values are influenced by the oxygen isotope composition of the food and water ingested by an animal as well as by certain physiological processes (such as sweating, panting and respiration) lower than those for terrestrial mammals
lower than those for terrestrial mammals 18O and
18O and ![Figure 4 : Bivariate plot of |[dgr]|18O and |[dgr]|13C values for enamel samples of early and middle Eocene mammals from India and Pakistan. Unfortunately we are unable to provide accessible alternative text for this. If you require assistance to access this image, or to obtain a text description, please contact npg@nature.com](http://origin.www.nature.com/nature/journal/v450/n7173/images/nature06343-f4.0.jpg)







 Tomaso schrijft over
Tomaso schrijft over 





{kind=link}
{kind=link}